2.1 The Cells of the Nervous System
The nervous system is composed of two main classes of cells: neurons and glial cells. Neurons are the basic signaling units that transmit information throughout the nervous system. As Santiago Ramón y Cajal and others of his time deduced, neurons take in information, make a “decision” about it following some relatively simple rules, and then, by changes in their activity levels, pass the signal along to other neurons or muscles. Neurons vary in their form, location, and interconnectivity within the nervous system (Figure 2.2), and these variations are closely related to their functions. Glial cells serve various functions in the nervous system, providing structural support and electrical insulation to neurons and modulating neuronal activity. We begin with a quick look at glial cells, after which we will turn our focus back to neurons.
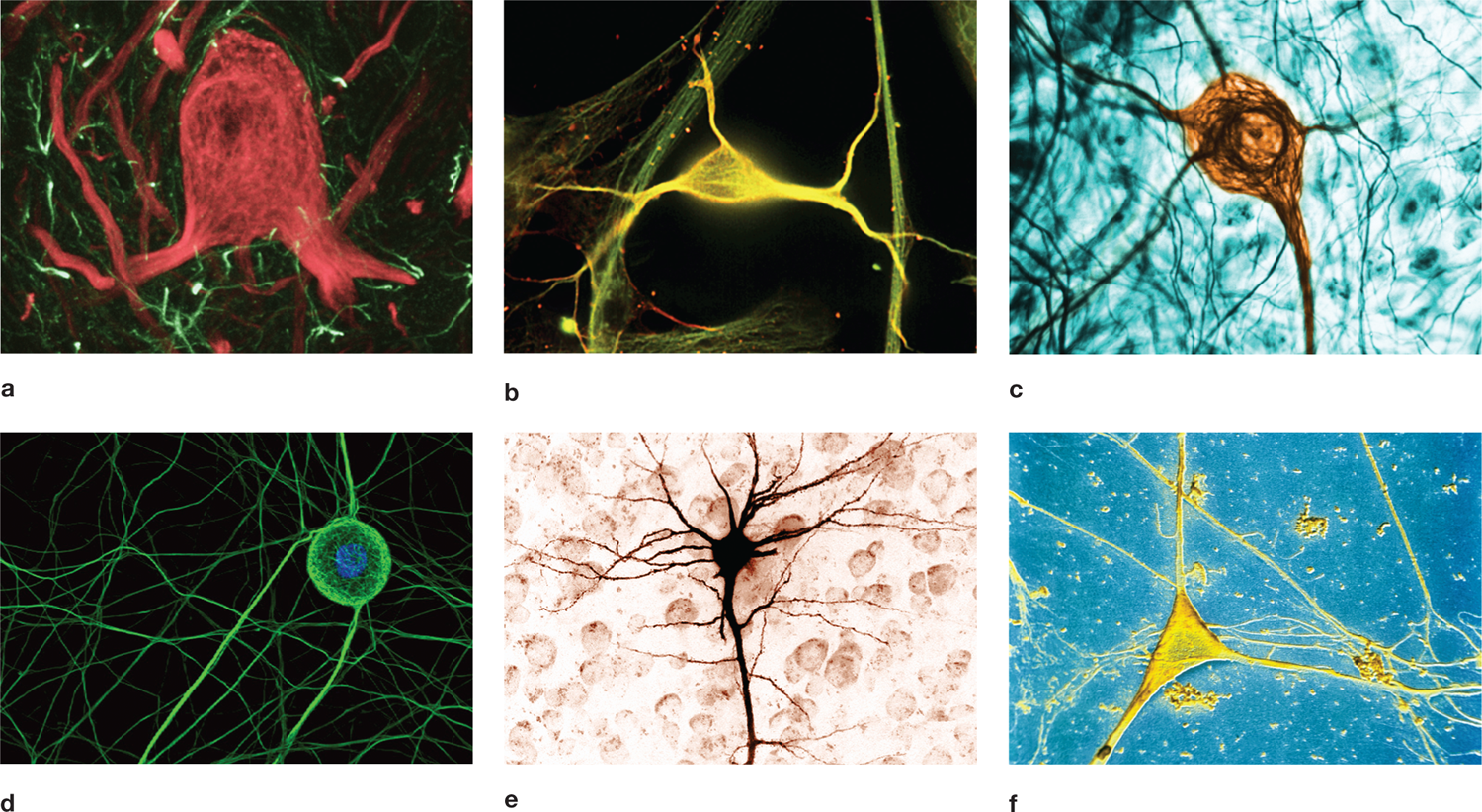
FIGURE 2.2 Mammalian neurons show enormous anatomical variety.
(a) Neuron (red) from the vestibular area of the brain. Glial cells are the thin, lighter structures (confocal light micrograph). (b) Hippocampal neuron (yellow; fluorescent micrograph). (c) Neuron (brown) in mouse dorsal root ganglion of the spinal cord (transmission electron micrograph). (d) Neuron in cell culture from dorsal root ganglia of an embryonic rat (fluorescent micrograph). (e) Pyramidal neuron from the brain. (f) Multipolar neuron cell body from human cerebral cortex (scanning electron micrograph).
Glial Cells
There are roughly as many glial cells in the brain as there are neurons. The central nervous system has three main types of glial cells: astrocytes, microglial cells, and oligodendrocytes (Figure 2.3). Astrocytes are large glial cells with round or radially symmetrical forms; they surround neurons and are in close contact with the brain’s vasculature. An astrocyte makes contact with blood vessels at specializations called end feet, which permit the astrocyte to transport ions across the vascular wall.
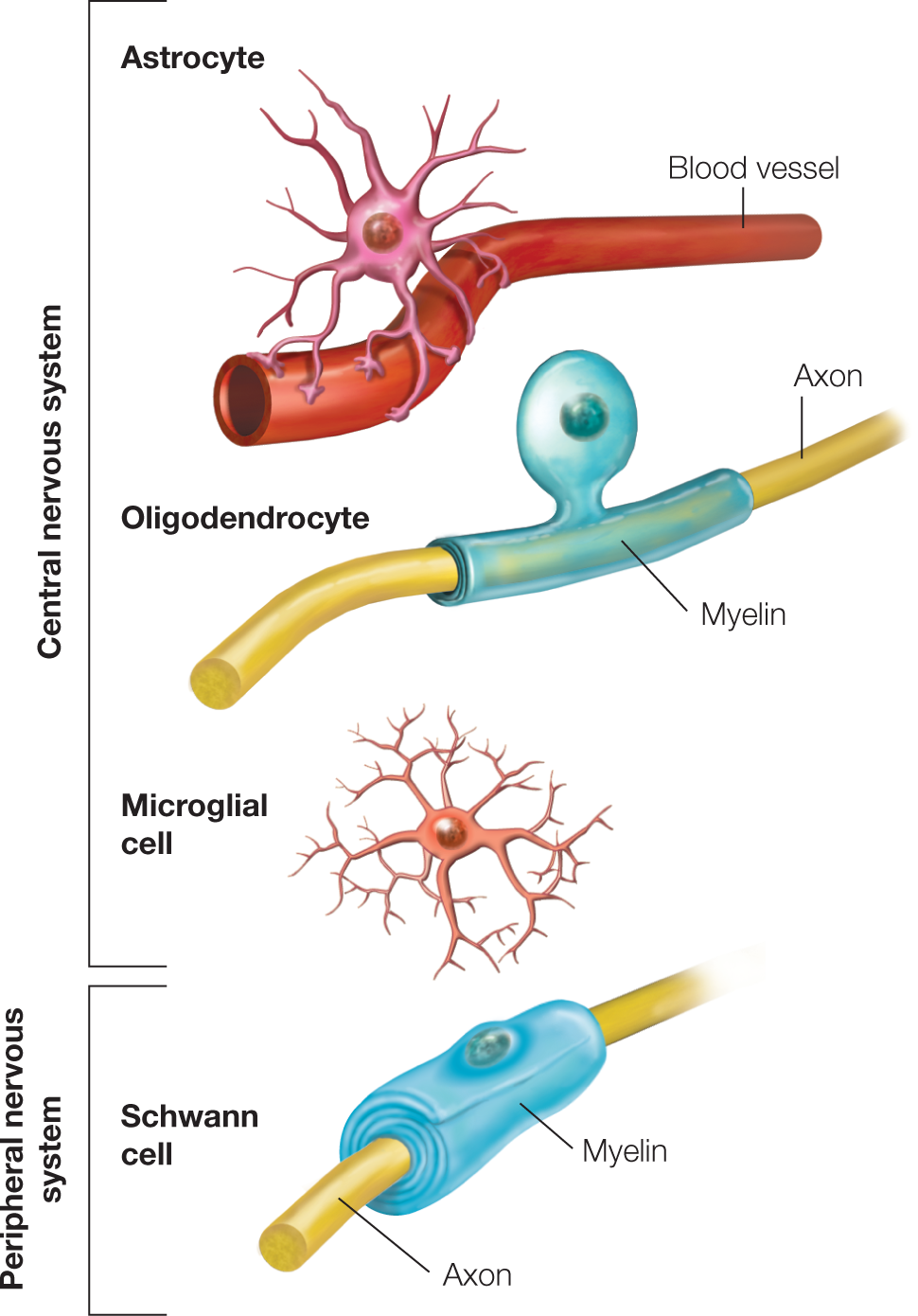
FIGURE 2.3 Various types of glial cells in the mammalian central and peripheral nervous systems.
An astrocyte is shown with end feet attached to a blood vessel. Oligodendrocytes and Schwann cells produce myelin around the axons of neurons (oligodendrocytes in the central nervous system, Schwann cells in the peripheral nervous system). Microglial cells dispose of damaged cells.
The astrocytes create a barrier, called the blood–brain barrier (BBB), between the tissues of the central nervous system and the blood. The BBB restricts the diffusion of microscopic objects (such as most bacteria) and large hydrophilic molecules in the blood from entering the neural tissue, but it allows the diffusion of small hydrophobic molecules, such as oxygen, carbon dioxide, and hormones. Many drugs and certain neuroactive agents, including dopamine and norepinephrine, when placed in the blood, cannot cross the BBB. Thus, it plays a vital role in protecting the central nervous system from blood-borne agents such as chemical compounds and also pathogens.
Evidence gathered over the past decade suggests that astrocytes also have an active role in brain function. In vitro studies indicate that they respond to and release neurotransmitters and other neuroactive substances that affect neuronal activity and modulate synaptic strength. In vivo studies found that when astrocyte activity is blocked, neuronal activity increases, supporting the notion that neuronal activity is moderated by astrocyte activity (Schummers et al., 2008). It is hypothesized that astrocytes either directly or indirectly regulate the reuptake of neurotransmitters.
Glial cells also form the fatty substance called myelin in the nervous system. In the central nervous system, oligodendrocytes form myelin; in the peripheral nervous system, Schwann cells carry out this task (Figure 2.3). Both glial cell types create myelin by wrapping their cell membranes around the axon in a concentric manner during development and maturation. The cytoplasm in that portion of the glial cell is squeezed out, leaving layers of the lipid bilayer of the glial cell sheathing the membrane. Myelin is a good electrical insulator, preventing loss of electrical current across the cell membrane. It increases the speed and distance that information can travel along a neuron.
Microglial cells, which are small and irregularly shaped (Figure 2.3), are phagocytes that devour and remove damaged cells. Unlike many cells in the central nervous system, microglial cells can proliferate even in adults (as do other glial cells).
Neurons
The standard cellular components found in almost all eukaryotic cells are also found in neurons (as well as in glial cells). A cell membrane encases the cell body (in neurons, it is sometimes called the soma; Greek for “body”), which contains the metabolic machinery that maintains the neuron: nucleus, endoplasmic reticulum, cytoskeleton, mitochondria, Golgi apparatus, and other common intracellular organelles (Figure 2.4). These structures are suspended in cytoplasm, the salty intracellular fluid that is made up of a combination of ions (molecules or atoms that have either a positive or negative electrical charge)—predominantly ions of potassium, sodium, chloride, and calcium—as well as molecules such as proteins. The neuron itself, like any other cell, sits in a bath of salty extracellular fluid, which is made up of a mixture of the same types of ions.
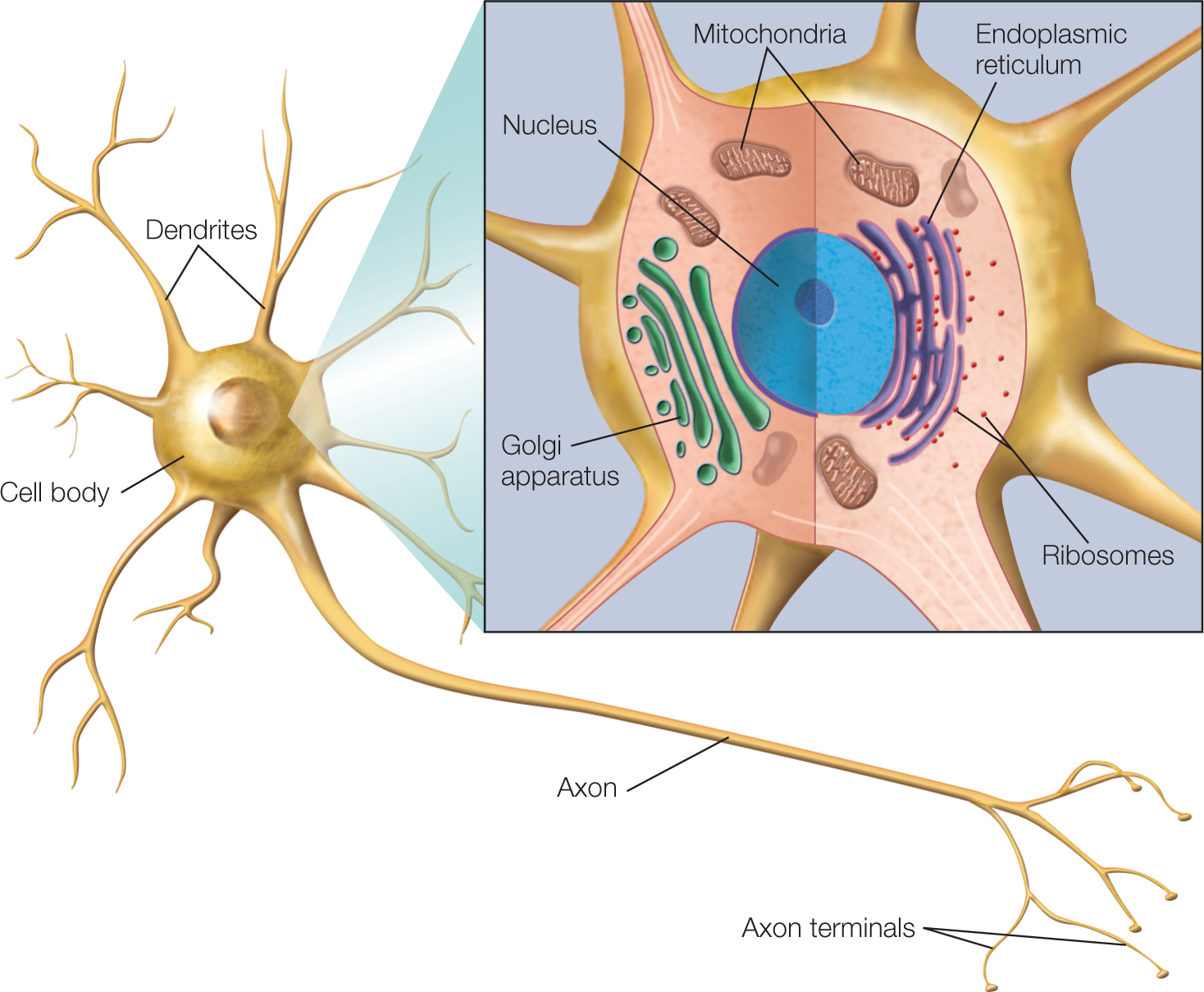
FIGURE 2.4 Idealized mammalian neuron.
A neuron is composed of three main parts: a cell body, dendrites, and an axon. The cell body contains the cellular machinery for the production of proteins and other macromolecules. Like other cells, the neuron contains a nucleus, endoplasmic reticulum, ribosomes, mitochondria, Golgi apparatus, and other intracellular organelles (inset). The dendrites and axon are extensions of the cell membrane and contain cytoplasm continuous with the cytoplasm inside the cell body.
In addition, however, neurons possess unique cytological features and physiological properties that enable them to transmit and process information rapidly. The two predominant cellular components unique to neurons are the dendrites and the axon. Dendrites are branching extensions of the neuron that receive inputs from other neurons. They have many varied and complex forms, depending on the type and location of the neuron. The arborizations may look like the branches and twigs of an oak tree, as seen in the complex dendritic structures of the cerebellar Purkinje cells (Figure 2.5), or they may be much simpler, such as the dendrites in spinal motor neurons (Figure 2.6). Most dendrites also have specialized processes called spines, little knobs attached by small necks to the surface of the dendrites, where the dendrites receive inputs from other neurons (Figure 2.7).
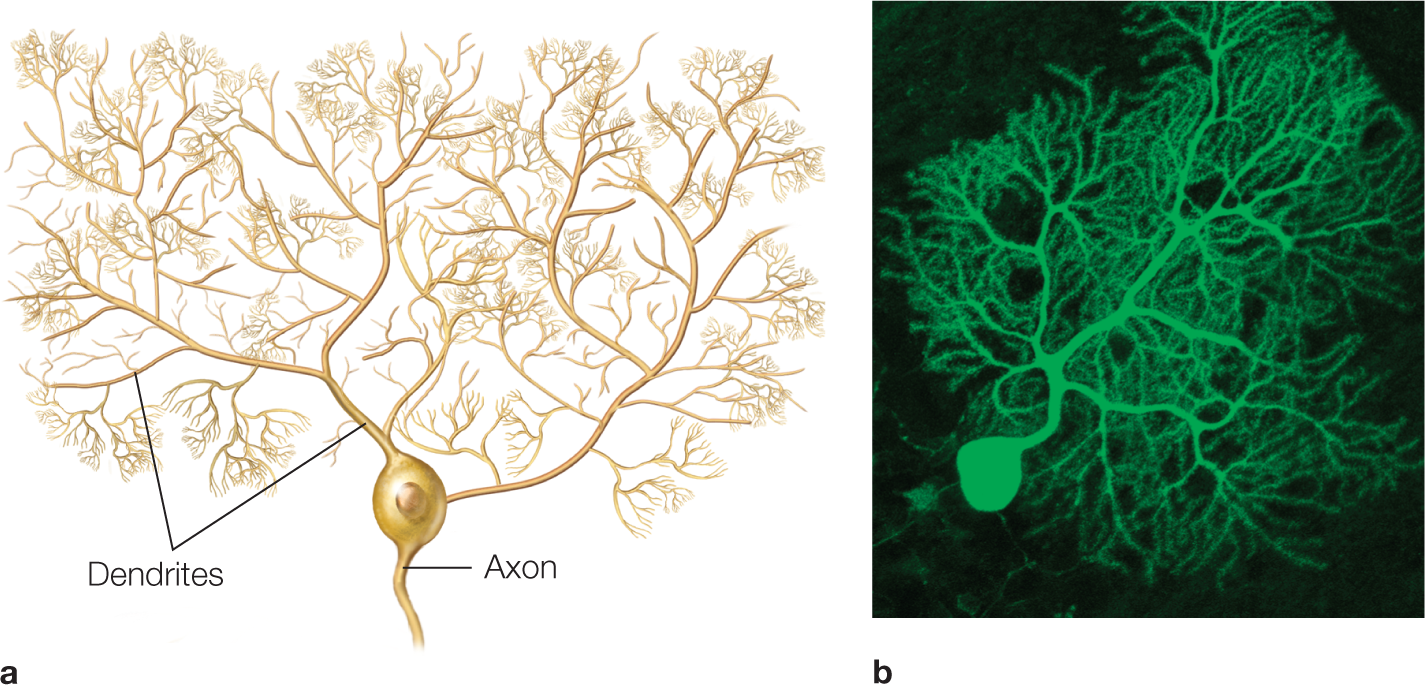
FIGURE 2.5 Soma and dendritic tree of a Purkinje cell from the cerebellum.
The Purkinje cells are arrayed in rows in the cerebellum. Each one has a large dendritic tree that is wider in one direction than the other. (a) Drawing of Purkinje cell as viewed in a cross section through a cerebellar folium. (b) Confocal micrograph of a Purkinje cell from mouse cerebellum. The cell is visualized using fluorescence methods.
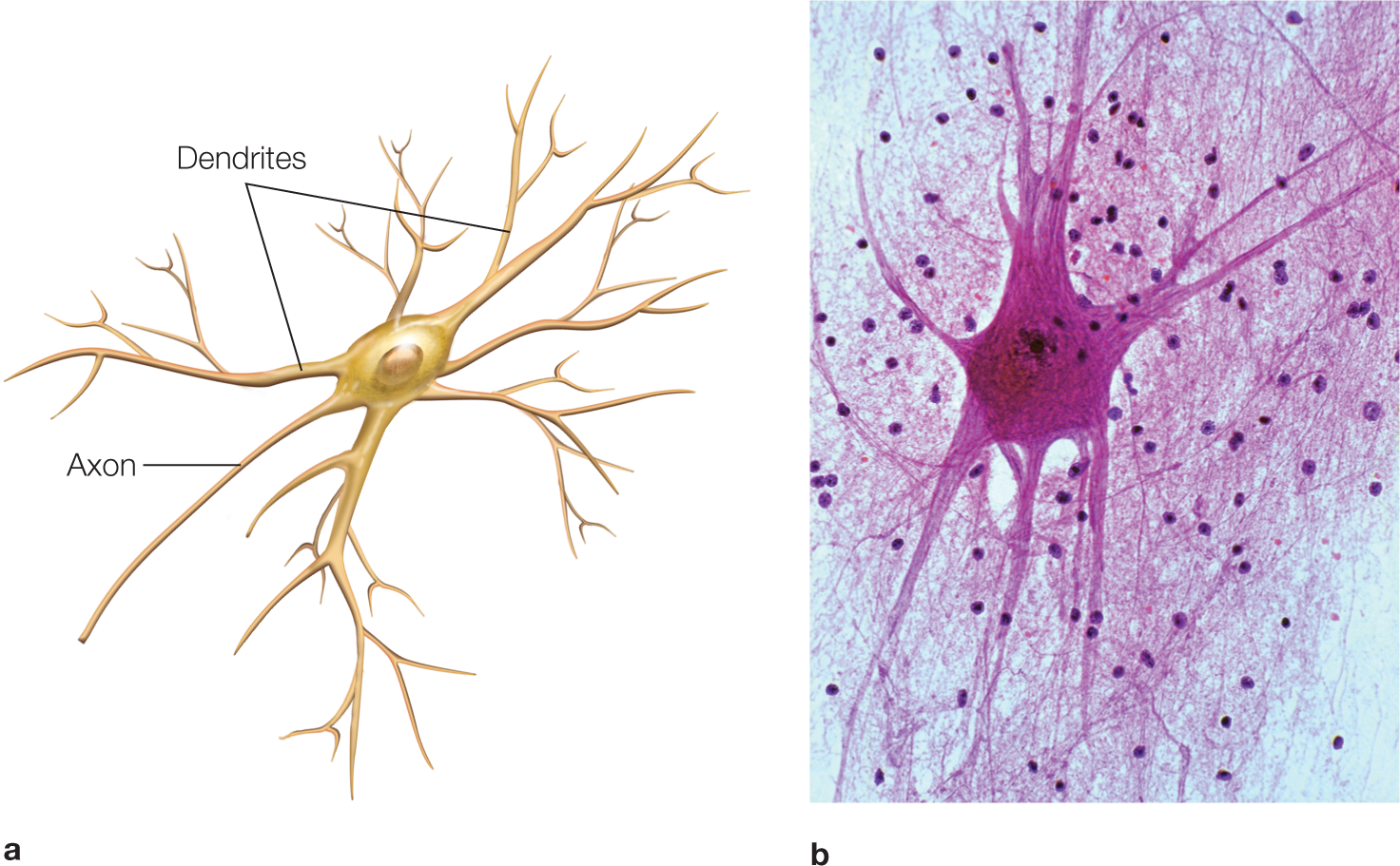
FIGURE 2.6 Spinal motor neuron.
(a) Neurons located in the ventral horn of the spinal cord send their axons out the ventral root to make synapses on muscle fibers. (b) A spinal cord motor neuron stained with cresyl echt violet stain.
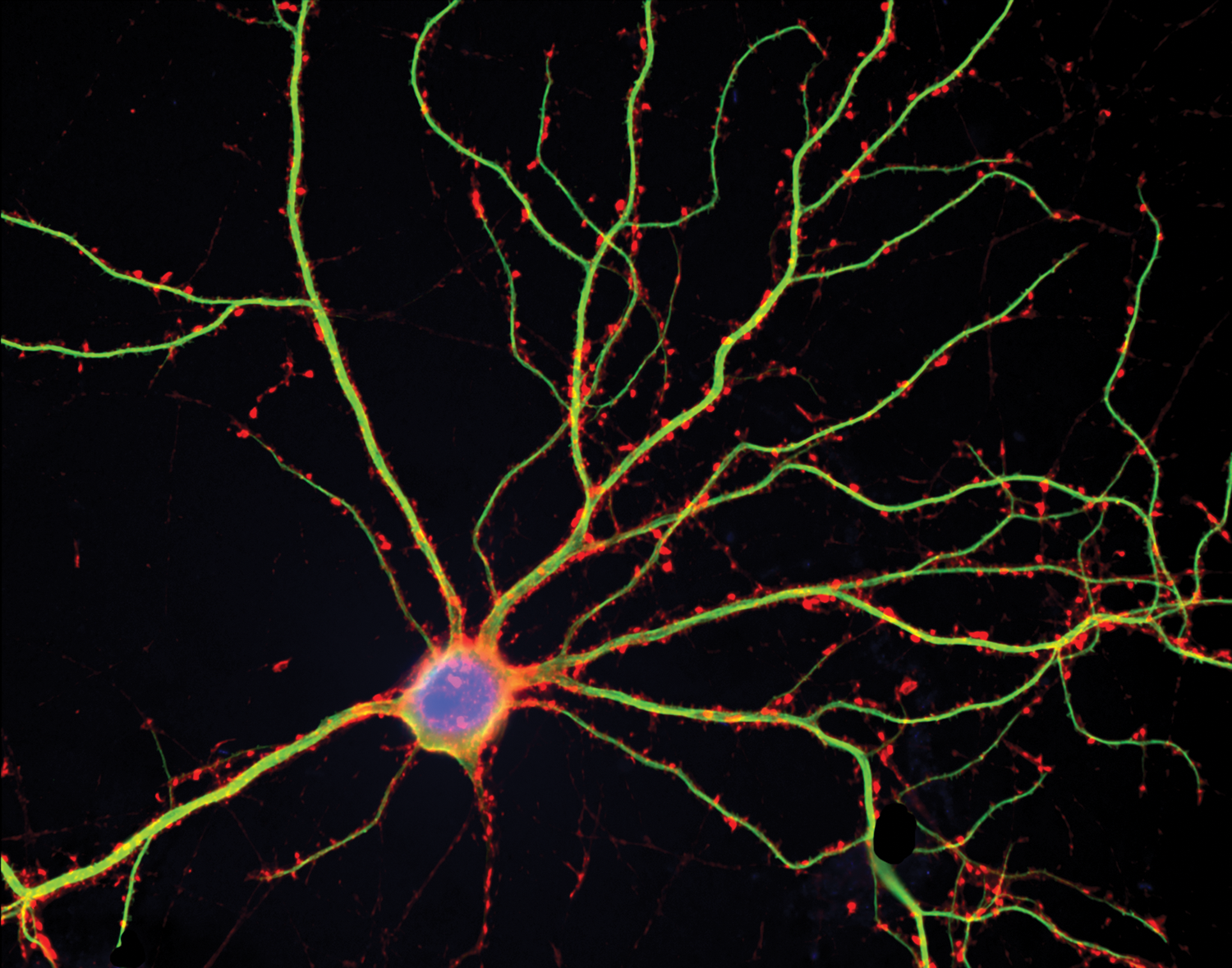
FIGURE 2.7 Dendritic spines on cultured rat hippocampal neurons.
This neuron has been triple stained to reveal the cell body (blue), dendrites (green), and spines (red).
The axon is a single process that extends from the cell body. This structure represents the output side of the neuron. Electrical signals travel along the length of the axon to its end, the axon terminals, where the neuron transmits the signal to other neurons or other targets. Transmission occurs at the synapse, a specialized structure where two neurons come into close contact so that chemical or electrical signals can be passed from one cell to the next. Some axons branch to form axon collaterals that can transmit signals to more than one cell (Figure 2.8).
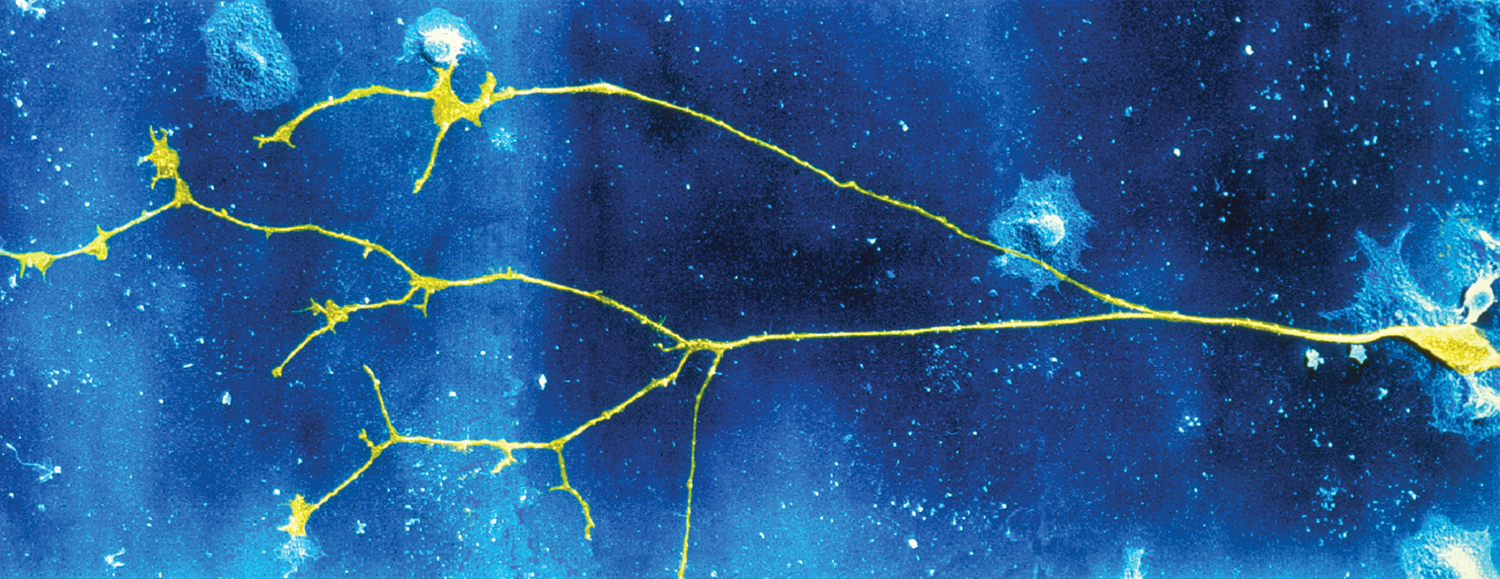
FIGURE 2.8 Axons can take different forms.
A neuron (far right) and its axon collaterals (left half of image) are shown stained in yellow. The cell body (at far right) gives rise to an axon, which branches, forming collaterals that can make contact with many different neurons.
Many axons are wrapped in layers of myelin. Along the length of the axons, there are evenly spaced gaps in the myelin; these gaps are commonly referred to as nodes of Ranvier (see Figure 2.12). Later, when we look at how signals move down an axon, we will explore the role of myelin and the nodes of Ranvier in accelerating signal transmission.
Neuronal Signaling
Neurons receive, evaluate, and transmit information. These processes are referred to as neuronal signaling. Information that is received by the neuron at its input synapses passes through the cell body and then, via the axon, to output synapses on the axon terminals. At these output synapses, information is transferred across synapses from one neuron to the next neuron; or to nonneuronal cells such as those in muscles or glands; or to other targets, such as blood vessels.
Within a neuron, information moves from input synapses to output synapses through changes in the electrical state of the neuron caused by the flow of electrical currents within the neuron and across its neuronal membrane. Between neurons, information transfer across synapses is typically mediated chemically by neurotransmitters (signaling molecules); these synapses are called chemical synapses. At electrical synapses, however, signals between neurons travel via transsynaptic electrical currents. Regarding information flow, neurons are referred to as either presynaptic or postsynaptic in relation to any particular synapse. Most neurons are both presynaptic and postsynaptic: They are presynaptic when their axon’s output synapses make connections onto other neurons or targets, and they are postsynaptic when other neurons make a connection at input synapses onto their dendrites or elsewhere on the receiving neuron.
The Membrane Potential The process of signaling has several stages. Let’s return to Delgado’s bull, whose neurons process information in the same way ours do. The bull is snorting about in the dirt, head down, when suddenly a sound wave—produced by Delgado entering the ring—courses down its auditory canal and hits the tympanic membrane (eardrum). The resultant stimulation of the auditory receptor cells (auditory hair cells) generates neuronal signals that are transmitted via the auditory pathways to the brain. At each stage of this ascending auditory pathway, neurons receive inputs on their dendrites that typically cause them to generate signals that are transmitted to the next neuron in the pathway.
How does the neuron generate these signals, and what are these signals? To answer these questions, we have to understand several things about neurons. First, energy is needed to generate the signals. Second, this energy is in the form of an electrical potential across the neuronal membrane. This electrical potential is defined as the difference in voltage across the neuronal membrane or, put simply, the voltage inside the neuron versus outside the neuron. Third, these two voltages depend on the concentrations of potassium, sodium, and chloride ions, as well as on charged protein molecules both inside and outside of the cell. Fourth, when a neuron is in its resting state and not actively signaling, the inside of a neuron is more negatively charged than the outside. The voltage difference across the neuronal membrane in the resting state is typically about –70 millivolts (mV) inside, which is known as the resting potential or resting membrane potential. This electrical-potential difference means that the neuron has at its disposal a kind of battery; and like a battery, the stored energy can be used to do work—signaling work (Figure 2.9).
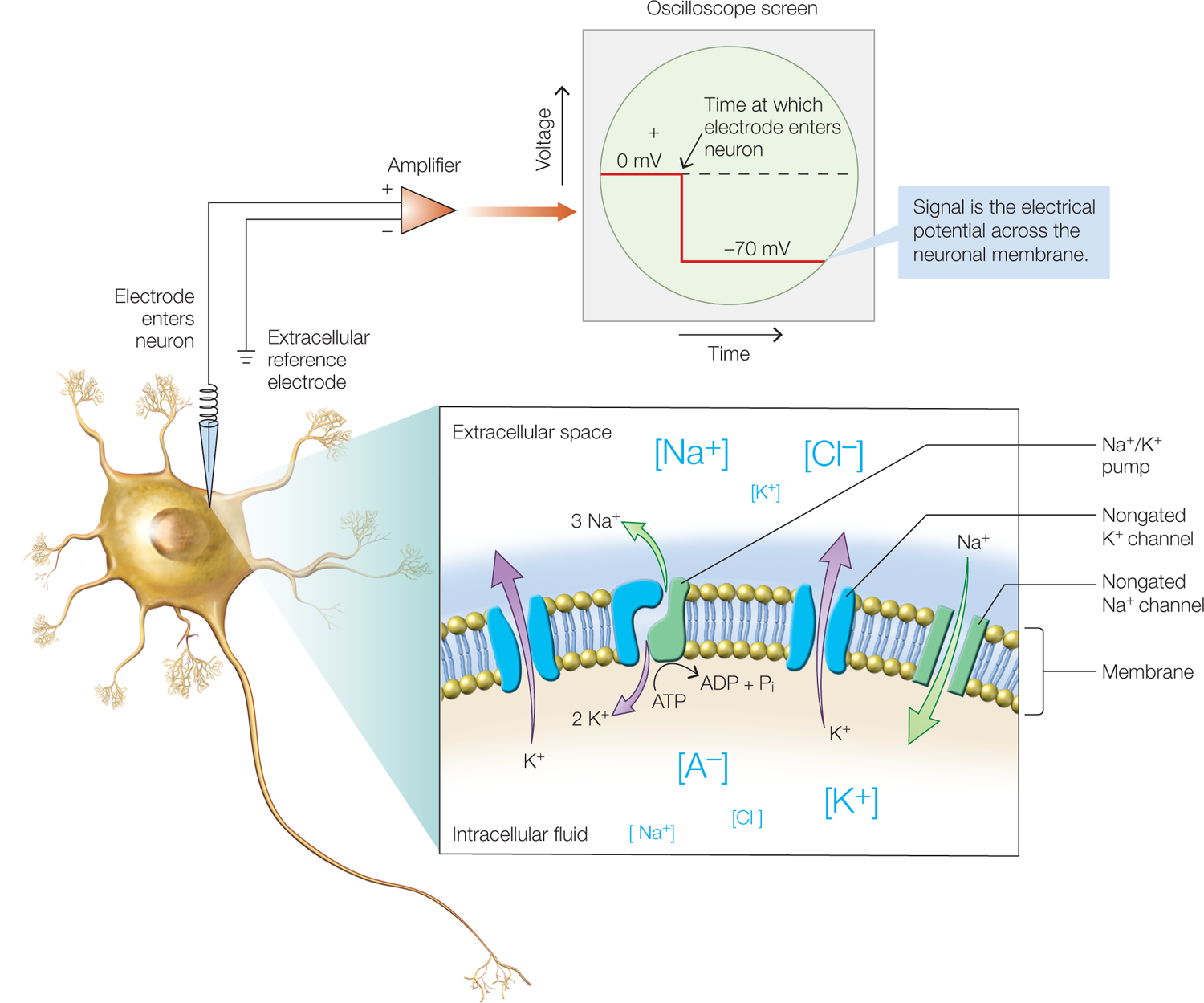
FIGURE 2.9 Ion channels in a segment of neuronal membrane and measuring resting membrane potential.
Idealized neuron (left), shown with an intracellular recording electrode penetrating the neuron. The electrode measures the difference between the voltage inside versus outside the neuron, and this difference is amplified and displayed on an oscilloscope screen (top). The oscilloscope screen shows voltage over time. Before the electrode enters the neuron, the voltage difference between the electrode and the extracellular reference electrode is zero, but when the electrode is pushed into the neuron, the difference becomes –70 mV, which is the resting membrane potential. The resting membrane potential arises from the asymmetrical distribution of ions of sodium (Na+), potassium (K+), and chloride (Cl-), as well as of charged protein molecules (A-), across the neuron’s cell membrane (inset).
How does the neuron generate and maintain this resting potential, and how does it use it for signaling? To answer these questions about function, we first need to examine the structures in the neuron that are involved in signaling. The bulk of the neuronal membrane is a bilayer of fatty lipid molecules that separates the cytoplasm from the extracellular milieu. Because the membrane is composed of lipids, it does not dissolve in the watery environments found inside and outside of the neuron, and it blocks the flow of water-soluble substances between the inside and the outside. It prevents ions, proteins, and other water-soluble molecules from moving across it. To understand neuronal signaling, we must focus on ions. This point is important: The lipid membrane maintains the separation of intracellular and extracellular ions and electrical charge that ultimately permits neuronal communication.
The neuronal membrane, though, is not merely a lipid bilayer. The membrane is peppered with transmembrane proteins, some of which serve as conduits for ions to move across the membrane (Figure 2.9, inset). These proteins are of two main types: ion channels and ion pumps. Ion channels, as we will see, are proteins with a pore through the center, and they allow certain ions to flow down their electrochemical and concentration gradients. Ion pumps use energy to actively transport ions across the membrane against their concentration gradients—that is, from regions of low concentration to regions of higher concentration.
Ion channels. The transmembrane passageways created by ion channels are formed from the three-dimensional structure of these proteins. These hydrophilic channels selectively permit one type of ion to pass through the membrane. The ion channels of concern to us—the ones found in neurons—are selective for sodium, potassium, calcium, or chloride ions (Na+, K+, Ca2+, and Cl-, respectively; Figure 2.9, inset). The extent to which a particular ion can cross the membrane through a given ion channel is referred to as its permeability. This characteristic of ion channels gives the neuronal membrane the attribute of selective permeability. (Selective permeability is actually a property of all cells in the body; as part of cellular homeostasis, it enables cells to maintain internal chemical stability.) The neuronal membrane is more permeable to K+ than to Na+ (or other) ions—a property that contributes to the resting membrane potential, as we will learn shortly. The membrane permeability to K+ is larger because there are many more K+-selective channels than any other type of ion channel.
Unlike most cells in the body, neurons are excitable, meaning that their membrane permeability can change (because the membranes have ion channels that are capable of changing their permeability for a particular ion). Such proteins are called gated ion channels. They open or close in response to changes in nearby transmembrane voltage, or to chemical or physical stimuli. In contrast, ion channels that are unregulated, and hence always allow the associated ion to pass through, are known as nongated ion channels.
Ion pumps. Under normal conditions, Na+ and Cl- concentrations are greater outside the cell, and K+ concentrations are greater inside the cell. You may be wondering why K+ ions don’t flow out of the neuron—down their concentration gradient—until the K+ ion concentrations inside and outside the cell are equal. We could ask the corresponding question for all other ions.
To combat this drive toward equilibrium, neurons use active transport proteins known as ion pumps. In particular, neurons use a Na+/K+ pump that pumps Na+ ions out of the cell and K+ ions into the cell (Figure 2.9, inset). Because this process moves ions up their concentration gradients, the mechanism requires energy. Each pump is an enzyme that hydrolyzes adenosine triphosphate (ATP) for energy. For each molecule of ATP that is hydrolyzed, enough energy is produced to move three Na+ ions out of the cell and two K+ ions into the cell (Figure 2.10).
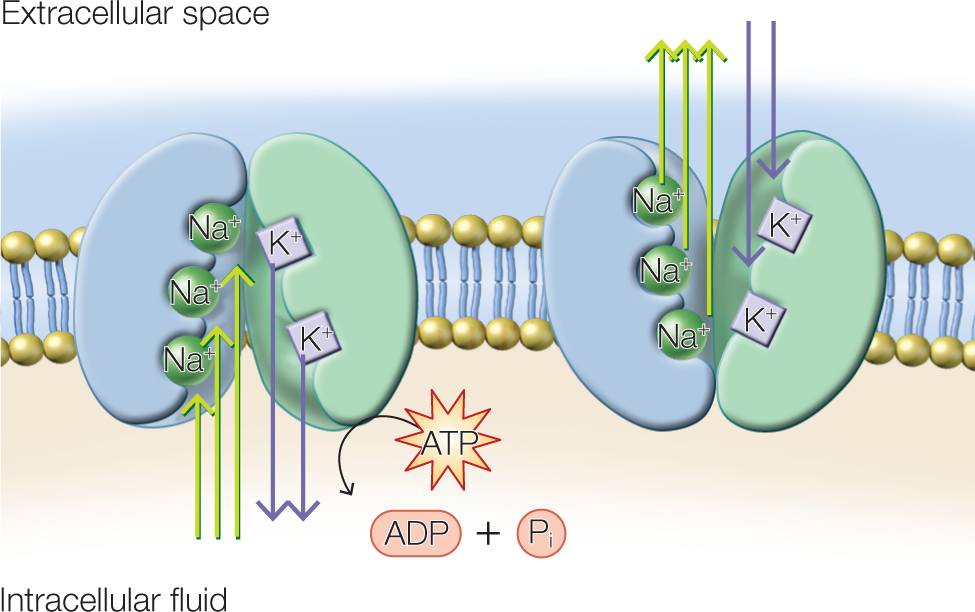
FIGURE 2.10 Ion pumps transport ions across the membrane.
The Na+/K+ pump preserves the cell’s resting potential by maintaining a larger concentration of K+ inside the cell and Na+ outside the cell. The pump uses ATP as energy.
The concentration gradients create forces due to the unequal distribution of ions. The force of the Na+ concentration gradient acts to push Na+ from an area of high concentration to one of low concentration (from outside to inside), while the K+ concentration gradient acts to push K+ also from an area of high concentration to an area of low concentration (from inside to outside)—the very thing the pump is working against. With both positively and negatively charged ions inside and outside the cell, why is the voltage different inside versus outside the neuron?
The inside and outside voltages are different because the membrane is more permeable to K+ than to Na+. The force of the K+ concentration gradient pushes some K+ out of the cell, leaving the inside of the neuron slightly more negative than the outside. This difference creates another force, an electrical gradient, because each K+ ion carries one unit of positive charge out of the neuron as it moves across the membrane. These two gradients (electrical and ionic concentration) are in opposition to one another with respect to K+ (Figure 2.11).
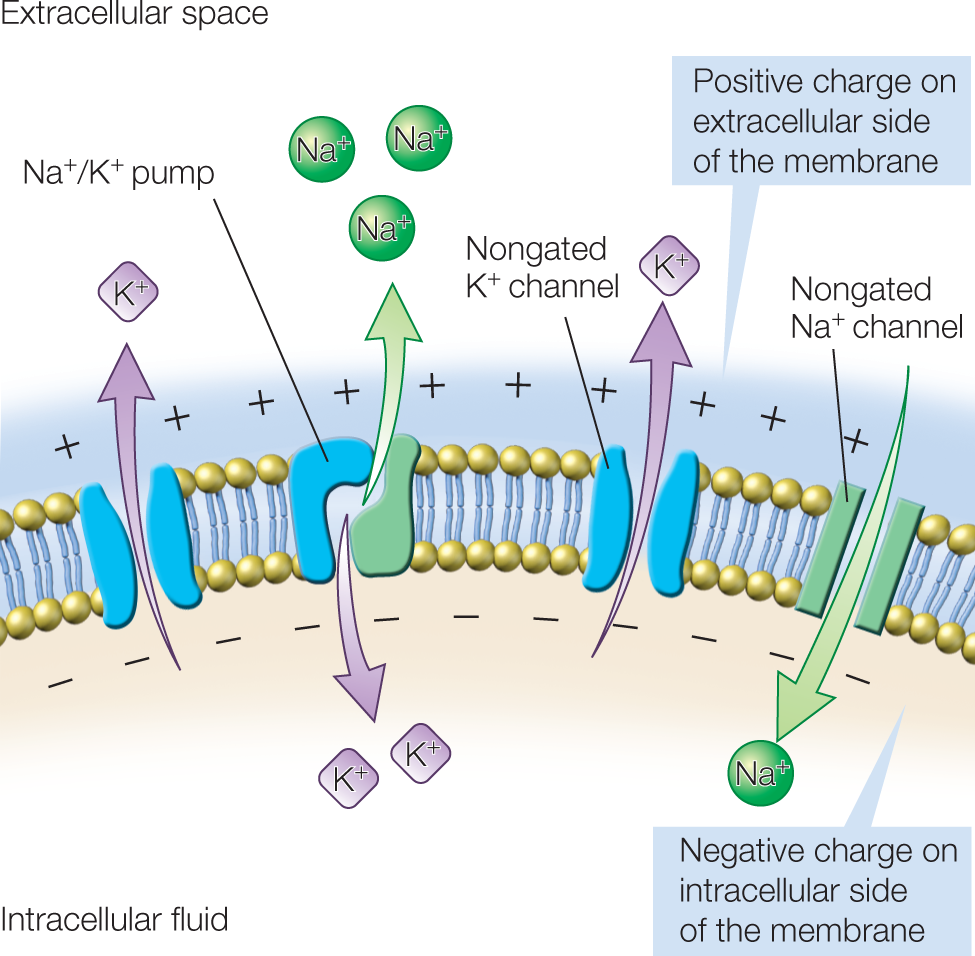
FIGURE 2.11 Selective permeability of the membrane.
The membrane’s selective permeability to some ions, and the concentration gradients formed by active pumping, lead to a difference in electrical potential across the membrane; this is the resting membrane potential. The membrane potential, represented here by the positive charges outside the neuron along the membrane and the negative charges inside along the membrane, is the basis for the transmembrane voltage difference shown in Figure 2.9.
As negative charge builds up along the inside of the membrane (and an equivalent positive charge forms along the extracellular side), the positively charged K+ ions outside of the cell are drawn electrically back into the neuron through the same ion channels that are allowing K+ ions to leave the cell by diffusion. Eventually, the force of the concentration gradient pushing K+ out through the K+ channels is equal to the force of the electrical gradient driving K+ in. When that happens, the opposing forces are said to reach electrochemical equilibrium. The difference in charge thus produced across the membrane is the resting membrane potential, that −70 mV difference. The value for the resting membrane potential of any cell can be calculated by using knowledge from electrochemistry, provided that the concentrations of ions inside and outside the neuron are known.
The Action Potential We now understand the basis of the energy source that neurons can use for signaling. Next we want to learn how this energy can be used to transmit information within a neuron, from its dendrites, which receive inputs from other neurons, to its axon terminals, where it signals to the next neuron(s) in the chain. The process begins when synapses on a neuron’s dendrites receive a signal (e.g., a neurotransmitter binding to a receptor), resulting in the opening of ion channels in the dendrite, which causes ionic currents to flow. For excitatory synaptic inputs, excitatory postsynaptic potentials (EPSPs) occur in the dendrite, and ionic currents flow through the volume of the neuron’s cell body. If these currents happen to be strong enough to reach distant axon terminals, then the process of neuronal signaling is complete. In the vast majority of cases, however, the distance from dendrites to axon terminals is too great for the EPSP to have any effect. Why?
The small electrical current produced by the EPSP is passively conducted through the cytoplasm of the dendrite, cell body, and axon. Passive current conduction is called electrotonic conduction or decremental conduction: “decremental” because it diminishes with distance from its origin—the synapse on the dendrites, in this case. The maximum distance a passive current will flow in a neuron is only about 1 mm. In most neurons, a millimeter is too short to effectively conduct electrical signals, although sometimes, like in a structure such as the retina, a millimeter is sufficient to permit neuron-to-neuron communication via decremental conduction. Most of the time, however, the reduction in signal intensity with decremental conduction means that long-distance communication within a neuron from dendrite to axon terminal will fail (your toes would be in trouble, for example, because they are about 1 meter from the spinal cord and close to 2 meters from the brain). How does the neuron solve this problem of decremental conduction and the need to conduct signals over long distances?
Neurons evolved a clever mechanism to regenerate and pass along the signal received at synapses on the dendrite: the action potential. It works something like 19th-century firefighters in a bucket brigade. An action potential is a rapid depolarization and repolarization of a small region of the membrane on the neuron’s output via its axon caused by the opening and closing of ion channels.
An action potential is a different process from the EPSP and the ionic currents involved in decremental conduction. The action potential doesn’t decrement after only 1 mm. Action potentials enable signals to travel for meters with no loss in signal strength, because they continually regenerate the signal at each patch of membrane on the axon. This is one reason why giraffes and blue whales can have neurons whose axon terminals may be many meters from their dendrites.
The action potential is able to regenerate itself because of the presence of voltage-gated ion channels located in the neuronal membrane (Figure 2.12a, inset). The densest concentration of ion channels is found at the spike-triggering zone in the axon hillock, a specialized region of the neuronal soma where the axon emerges. As its name denotes, the spike-triggering zone initiates the action potential. (The term spike is shorthand for an action potential because it represents a rapid change and a peak in the depolarization of the membrane potential, and it actually looks like a spike when viewed as a recording displayed on an oscilloscope or computer screen.) Ion channels are also found along the axon. In myelinated axons, voltage-gated ion channels along the axon’s length are restricted to the nodes of Ranvier. Named after the French histologist and anatomist Louis-Antoine Ranvier, who first described them, they are regular intervals along the axon where gaps in myelination occur (Figure 2.12a).
How does the spike-triggering zone initiate an action potential? The passive electrical currents that are generated following EPSPs on multiple distant dendrites sum together at the axon hillock. This current flows across the neuronal membrane in the spike-triggering zone, depolarizing the membrane. If the depolarization is strong enough, meaning that the membrane moves from its resting potential of about −70 mV to a less negative value of approximately −55 mV, then an action potential is triggered. We refer to this depolarized membrane potential value as the threshold for initiating an action potential. Figure 2.12b illustrates an idealized action potential. The numbered circles in the figure correspond to the numbered events in the next paragraph. Each event alters a small region of the membrane’s permeability for Na+ and K+ as a result of the opening and closing of voltage-gated ion channels.
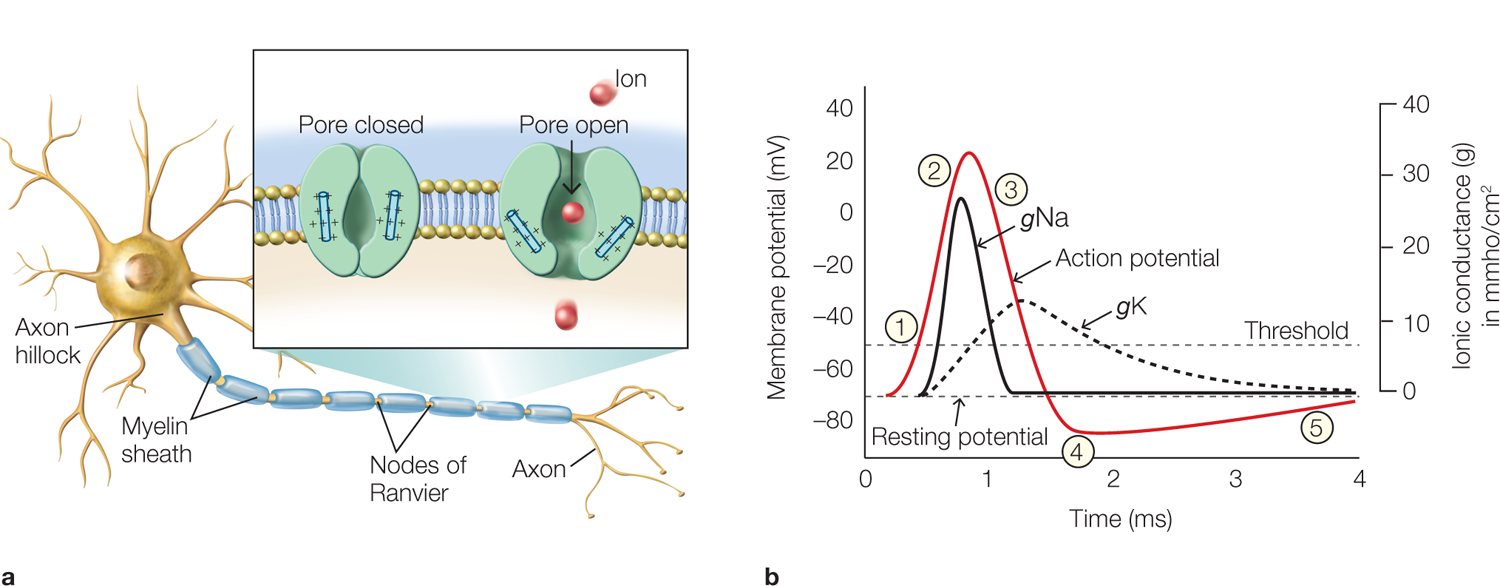
FIGURE 2.12 The neuronal action potential, voltage-gated ion channels, and changes in channel conductance.
(a) An idealized neuron with myelinated axon and axon terminals. Voltage-gated ion channels located in the spike-triggering zone at the axon hillock and along the extent of the axon at the nodes of Ranvier open and close rapidly, changing their conductance to specific ions (e.g., Na+), altering the membrane potential and resulting in the action potential (inset). (b) The relative time course of changes in membrane voltage during an action potential, and the underlying causative changes in membrane conductance to Na+ (gNa) and K+ (gK). The initial depolarizing phase of the action potential (red line) is mediated by increased Na+ conductance (black line), and the later repolarizing, descending phase of the action potential is mediated by an increase in K+ conductance (dashed line) that occurs when the K+ channels open. The Na1 channels have closed during the last part of the action potential, when repolarization by the K+ current is taking place. The action potential undershoots the resting membrane potential at the point where the membrane becomes more negative than the resting membrane potential.
When the threshold (Figure 2.12b, event 1) is reached, voltage-gated Na+ channels open and Na+ flows rapidly into the neuron. This influx of positive ions further depolarizes the neuron, opening additional voltage-gated Na+ channels; thus, the neuron becomes more depolarized (2), continuing the cycle by causing even more Na+ channels to open. This process is called the Hodgkin–Huxley cycle. This rapid, self-reinforcing cycle, lasting only about 1 ms, generates the large depolarization that is the first portion of the action potential. Next, the voltage-gated K+ channels open, allowing K+ to flow out of the neuron down its concentration gradient. This outward flow of positive ions begins to shift the membrane potential back toward its resting potential (3). The opening of the K+ channels outlasts the closing of the Na+ channels, causing a second repolarizing phase of the action potential; this repolarization drives the membrane potential toward the equilibrium potential of K+, which is even more negative than the resting potential. The equilibrium potential is the membrane potential at which there is no net flux of a given ion. As a result, the membrane is temporarily hyperpolarized: At about −80 mV, the membrane potential is more negative than both the resting membrane potential and the threshold required for triggering an action potential (4). Hyperpolarization causes the K+ channels to close, in response to which the membrane potential gradually returns to its resting state (5).
During this transient hyperpolarization state, the voltage-gated Na+ channels are unable to open, and no other action potential can be generated. This is known as the absolute refractory period. It is followed by the relative refractory period, during which the neuron can generate action potentials, but only with larger-than-normal depolarizing currents. The entire refractory period lasts only a couple of milliseconds and has two consequences. One is that the neuron’s speed for generating action potentials is limited to about 200 action potentials per second. The other is that the passive current that flows from the action potential cannot reopen the ion-gated channels that generated it. The passive current, however, does flow down the axon with enough strength to depolarize the membrane a bit farther on, where the ion channels are not in a refractory state, opening voltage-gated channels in this next portion of the membrane. The result is that the action potential moves down the axon in one direction only—from the axon hillock toward the axon terminal.
That is the story of the self-regenerating action potential as it propagates itself down an axon, sometimes traveling several meters in the process. But traveling far is not the end of the story. Action potentials must also travel quickly if a person wants to run, or a bull wants to charge, or a very large animal (think blue whale) simply wants to react in a reasonable amount of time. Accelerated transmission of the action potential is accomplished in myelinated axons. The thick lipid sheath of myelin (Figure 2.12a) surrounding the membrane of myelinated axons makes the axon super-resistant to voltage loss. The high electrical resistance allows passive currents generated by the action potential to be shunted farther down the axon. The result is that action potentials do not have to be generated as often, and they can be spread out along the axon at wider intervals.
Indeed, action potentials in myelinated axons need occur only at the nodes of Ranvier, where myelination is interrupted. As a result, the action potential appears to jump down the axon at great speed, from one node of Ranvier to the next. We call this saltatory conduction (the term is derived from the Latin word saltare, meaning “to jump or leap”). The importance of myelin for efficient neuronal conduction is notable when it is damaged or lost, which is what happens in multiple sclerosis (MS).
There is one more interesting tidbit concerning action potentials: Because they always have the same amplitude, they are said to be all-or-none phenomena. Since one action potential has the same amplitude as any other, the strength of the action potential does not communicate anything about the strength of the stimulus that initiated it. The intensity of a stimulus (e.g., a sensory signal) is communicated by the rate of firing of the action potentials: More intense stimuli elicit higher action-potential firing rates.
Neural Oscillations So far, we have presented an idealized situation: A neuron is sitting in a resting state, awaiting inputs that might cause it to experience EPSPs and action potentials at the axon hillock that move down the axon to transmit signals. But most neurons actually fire at a continuous baseline rate. This rate is different in different types of neurons, and it can be the result of either intrinsic properties of the neuron itself or activity in small neural circuits or larger neural networks. These neuronal oscillations are important for understanding some of the signals we can receive from electrodes placed in an intact brain or on the surface of the scalp. Because postsynaptic potentials like EPSPs can be recorded from populations of neurons, they are another measure of neuronal activity in addition to recordings of action potentials. In Chapter 3 we describe how these postsynaptic potentials, and not the action potentials, are the sources of electrical signals that can be recorded from the cortical surface or scalp in humans and animals using electroencephalography.
Take-Home Messages
- Glial cells form myelin around the axons of neurons. Myelin enables the rapid transmission of action potentials down an axon and increases the distance over which transmission can occur.
- Neurons communicate with other neurons and cells at specialized structures called synapses, where chemical and electrical signals can be conveyed between neurons.
- The electrical gradient across a neuron’s membrane results from the asymmetrical distribution of ions. The electrical difference across the membrane is the basis of the resting potential, the voltage difference across the neuronal membrane during rest (i.e., not during any phase of the action potential).
- Ion channels, formed by transmembrane proteins, can be either passive (always open) or gated (open only in the presence of electrical, chemical, or physical stimuli).
- Synaptic inputs result in postsynaptic potentials, and current to flow in the postsynaptic neuron.
- Postsynaptic currents can depolarize the axon hillock region, generating an action potential.
- Action potentials are all-or-none phenomena: The amplitude of the action potential does not depend on the size of the triggering depolarization, as long as that depolarization reaches the threshold for initiating the action potential.
- Nodes of Ranvier are the spaces between sheaths of myelin where voltage-gated Na+ and K+ channels are located and action potentials occur.
- Postsynaptic potentials lead to action potentials but also can be measured from large populations of neurons by electrodes located some distance away, such as the scalp, as when the oscillatory signals in an electroencephalogram (EEG) are being recorded.
- neuron
One of two cell types (along with the glial cell) in the nervous system. Neurons are responsible for processing sensory, motor, cognitive, and affective information.
- glial cell
One of two cell types (along with the neuron) in the nervous system. Glial cells are more numerous than neurons, by perhaps a factor of 10, and may account for more than half of the brain’s volume. They typically do not conduct signals themselves; but without them, the functionality of neurons would be severely diminished. Tissue made of glial cells is termed glia.
- blood–brain barrier (BBB)
A physical barrier formed by the end feet of astrocytes between the blood vessels in the brain and the tissues of the brain. The BBB limits which materials in the blood can gain access to neurons in the nervous system.
- myelin
A fatty substance that surrounds the axons of many neurons and increases the effective membrane resistance, helping to speed the conduction of action potentials.
- soma (pl. somata)
The cell body of a neuron.
- dendrites
Large treelike processes of neurons that receive inputs from other neurons at locations called synapses.
- spine
A little knob attached by a small neck to the surface of a dendrite. Synapses are located on spines.
- axon
The process extending away from a neuron down which action potentials travel. The terminals of axons contact other neurons at synapses.
- synapse
The specialized site on the neural membrane where a neuron comes close to another neuron to transmit information. Synapses include both presynaptic (e.g., synaptic vesicles with neurotransmitter) and postsynaptic (e.g., receptors) specializations in the neurons that are involved in chemical transmission. Electrical synapses involve special structures called gap junctions that make direct cytoplasmic connections between neurons.
- axon collaterals
Branches off an axon that can transmit signals to more than one cell.
- presynaptic
Referring to the neuron located before the synapse with respect to information flow. Compare postsynaptic.
- postsynaptic
Referring to the neuron located after the synapse with respect to information flow. Compare presynaptic.
- resting membrane potential
The difference in voltage across the neuronal membrane at rest, when the neuron is not signaling.
- ion channel
A passageway in the cell membrane, formed by a transmembrane protein that creates a pore, through which ions of a particular size and/or charge are allowed to pass.
- ion pump
A protein in the cell membrane of a neuron that is capable of transporting ions against their concentration gradient. The sodium–potassium pump transports sodium ions (Na+) out of the neuron and potassium ions (K+) into the neuron.
- permeability
The extent to which ions can cross a neuronal membrane.
- electrical gradient
A force that develops when a charge distribution across the neuronal membrane develops such that the charge inside is more positive or negative than the one outside. Electrical gradients result from asymmetrical distributions of ions across the membrane.
- electrotonic conduction
Also decremental conduction. Ionic current that flows passively through the cytoplasm and across the membrane of an activated neuron that diminishes with distance from the site of generation.
- action potential
The active or regenerative electrical signal that is required for synaptic communication. Action potentials are propagated along the axon and result in the release of neurotransmitters.
- voltage-gated ion channel
A transmembrane ion channel that changes molecular conformation when the membrane potential changes, altering the conductance of the channel for specific ions such as sodium, potassium, or chloride.
- spike-triggering zone
The location, at the juncture of the soma and the axon of a neuron, where currents from synaptic inputs on the soma and distant dendrites are summed and where voltage-gated Na+ channels are located that can be triggered to generate action potentials that can propagate down the axon.
- axon hillock
A part of the cell body of a neuron where the membrane potentials are summed before being transmitted down the axon.
- node of Ranvier
A location at which myelin is interrupted between successive patches of axon, and where an action potential can be generated.
- depolarization
A change in the membrane potential in which the electrical current inside the cell becomes less negative. With respect to the resting potential, a depolarized membrane potential is closer to the firing threshold. Compare hyperpolarization.
- threshold
The membrane potential value to which the membrane must be depolarized for an action potential to be initiated.
- equilibrium potential
The membrane potential at which a given ion (e.g., K+) has no net flux across the membrane—that is, the point where the numbers of ions moving outward and inward across the membrane are the same.
- hyperpolarization
A change in the membrane potential in which the electrical current inside of the cell becomes more negative. With respect to the resting potential, a hyperpolarized membrane potential is farther from the firing threshold. Compare depolarization.
- refractory period
The short period of time following an action potential during which the neuron may not be able to generate action potentials or may be able to do so only with larger-than-normal depolarizing currents.
- saltatory conduction
The mode of conduction in myelinated neurons, in which action potentials are generated down the axon only at nodes of Ranvier. Measurement of the propagation of the action potential gives it the appearance of jumping from node to node—hence the term saltatory, which comes from the Latin saltare, meaning “to jump.”