2.2 Synaptic Transmission
Neurons communicate with other neurons, with muscles, or with glands at synapses, and the transfer of a signal from the axon terminal of one neuron to the next neuron is called synaptic transmission. There are two major kinds of synapses—chemical and electrical—each using very different mechanisms for synaptic transmission.
Chemical Transmission
Most neurons send a signal to the cell across the synapse by releasing chemical neurotransmitters into the synaptic cleft, the gap between neurons at the synapse. The general mechanism is as follows: The arrival of the action potential at the axon terminal leads to depolarization of the terminal membrane, causing voltage-gated Ca2+ channels to open. The opening of these channels triggers small vesicles containing neurotransmitter to fuse with the membrane at the synapse and release the transmitter into the synaptic cleft. Different neurons produce and release different neurotransmitters, and some may release more than one type at a time, in what is called co-transmission. The transmitter diffuses across the cleft and, on reaching the postsynaptic membrane, binds with specific receptors embedded in it (Figure 2.13).
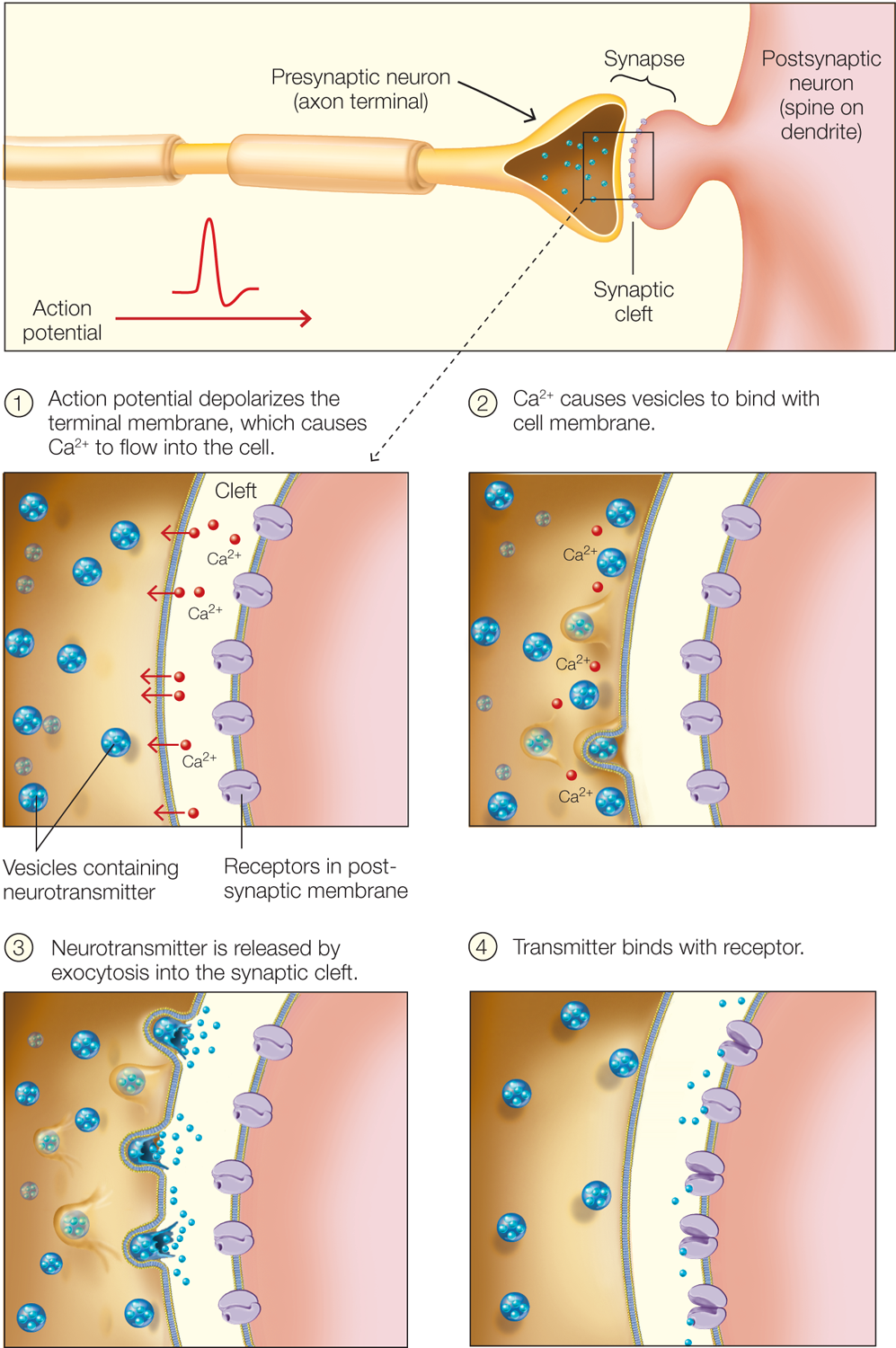
FIGURE 2.13 Neurotransmitter release at the synapse, into the synaptic cleft.
The synapse consists of various specializations where the presynaptic and postsynaptic membranes are close together. When the action potential invades the axon terminals, it causes voltage-gated Ca2+ channels to open (1), triggering vesicles to bind to the presynaptic membrane (2). Neurotransmitter is released into the synaptic cleft by exocytosis and diffuses across the cleft (3). Binding of the neurotransmitter to receptor molecules in the postsynaptic membrane completes the process of transmission (4).
There are two types of postsynaptic receptors: ligand-gated ion channels where neurotransmitter binding directly gates (opens) the ion channel, and G protein–coupled receptors (GPCRs) where biochemical signals indirectly cause the gating of ion channels; G proteins are those that bind the guanine nucleotides GDP and GTP (guanosine di- and triphosphate) and act as molecular switches in cells. Specific neurotransmitters bind to each type of postsynaptic receptor. In ligand-gated ion channels, binding induces a conformational change in the receptor. The change in shape opens an ion channel, resulting in an influx of ions leading to either depolarization (excitation) or hyperpolarization (inhibition) of the postsynaptic cell (Figure 2.14). Hyperpolarization of the postsynaptic neuron produces an inhibitory postsynaptic potential (IPSP).
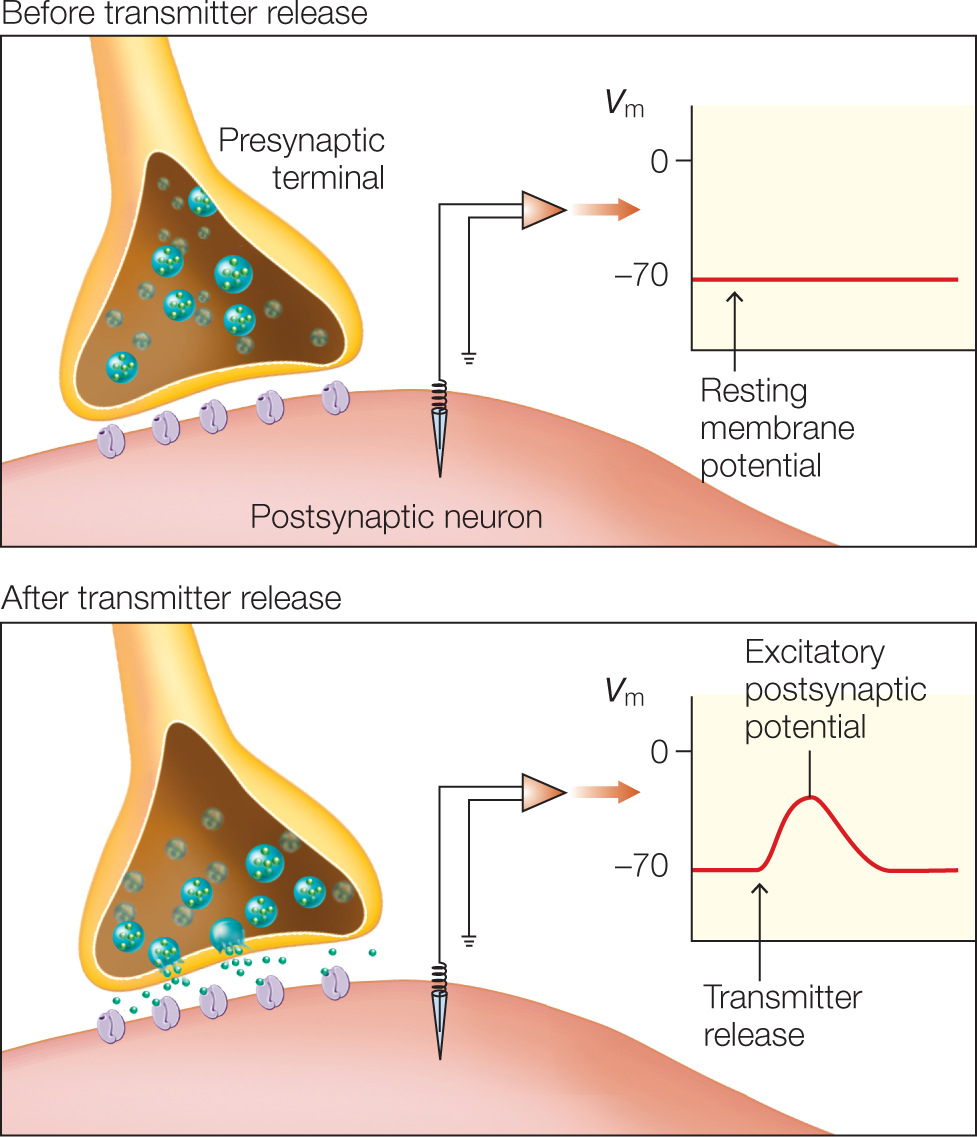
FIGURE 2.14 Neurotransmitter leading to a postsynaptic potential.
The binding of neurotransmitter to the postsynaptic membrane receptors changes the membrane potential (Vm). These postsynaptic potentials can be either excitatory (depolarizing the membrane), as shown here, or inhibitory (hyperpolarizing the membrane).
Excitatory and inhibitory neurons are also capable of modulating functions through the GPCRs. There are over 1,000 different GPCRs, giving us an idea of the complexity of the system. The particular GPCRs that are present depend on the neuron and where it is located. Each type of GPCR is activated by a specific signaling molecule, which could be a neurotransmitter, a neuropeptide (a small protein-like molecule secreted by neurons or glial cells), or a neurosteroid, among other possible signals.
When a signaling molecule specifically binds to its GPCR, the conformational change activates a G protein within the cell, which in turn activates or regulates a specific target protein, typically an enzyme, which produces a diffusible molecule of some sort called a second messenger. The second messenger, in turn, triggers a biochemical cascade of reactions. While directly gated channels mediate fast signaling, measured in milliseconds, GPCR-mediated signaling is slower, occurring over hundreds of milliseconds or even seconds and producing longer-lasting modulatory changes to the functional state. For example, the neurotransmitter epinephrine binds to a particular GPCR. Once bound, a G protein is activated that seeks out the protein adenylate cyclase and activates it. Activated adenylate cyclase turns ATP into cAMP (cyclic adenosine monophosphate), which acts as a second messenger of information instructing the postsynaptic neuron.
Neurotransmitters While you may have heard of a few of the classic neurotransmitters, more than 100 have been identified. What makes a molecule a neurotransmitter?
- It is synthesized by and localized within the presynaptic neuron, and stored in the presynaptic terminal before release.
- It is released by the presynaptic neuron when action potentials depolarize the terminal (mediated primarily by Ca2+).
- The postsynaptic neuron contains receptors specific for it.
- When artificially applied to a postsynaptic cell, it elicits the same response that stimulating the presynaptic neuron would.
Biochemical classification of neurotransmitters. Some neurotransmitters are amino acids: aspartate, gamma-aminobutyric acid (GABA), glutamate, and glycine. Another category of neurotransmitters, called biogenic amines, includes dopamine, norepinephrine, and epinephrine (these three are known as the catecholamines), serotonin (5-hydroxytryptamine), and histamine. Acetylcholine (ACh) is a well-studied neurotransmitter that is in its own biochemical class. Another large group of neurotransmitters consists of slightly larger molecules, the neuropeptides, which are made up of strings of amino acids. More than 100 neuropeptides are active in the mammalian brain, and they are divided into five groups:
- Tachykinins (brain-gut peptides, which are peptides secreted by endocrine cells and enteric neurons in the GI tract and also neurons in the central nervous system). This group includes substance P, which affects vasoconstriction and is a spinal neurotransmitter involved in pain.
- Neurohypophyseal hormones. Oxytocin and vasopressin are in this group. The former is involved in mammary functions and has been tagged the “love hormone” for its role in pair bonding and maternal behaviors; the latter is an antidiuretic hormone.
- Hypothalamic releasing hormones. This group includes corticotropin-releasing hormone, involved in the stress response; somatostatin, an inhibitor of growth hormone; and gonadotropin-releasing hormone, involved with the development, growth, and functioning of the body’s reproductive processes.
- Opioid peptides. This group is named for its similarity to opiate drugs, and these peptides bind to opiate receptors. It includes the endorphins and enkephalins.
- Other neuropeptides. This group includes peptides that do not fit neatly into another category, such as insulins, secretins (e.g., glucagon), and gastrins.
Some neurons produce only one type of neurotransmitter, but others produce multiple kinds. Neurons that do produce particular neurotransmitters sometimes form distinct systems, such as the cholinergic system, the noradrenergic system, the dopaminergic system, and the serotonergic system. When a neurotransmitter system is activated, large areas of the brain can be affected (Figure 2.15). Neurons that produce more than one type of transmitter may release them together or separately, depending on the conditions of stimulation. For example, the rate of stimulation by the action potential can induce the release of a specific neurotransmitter.
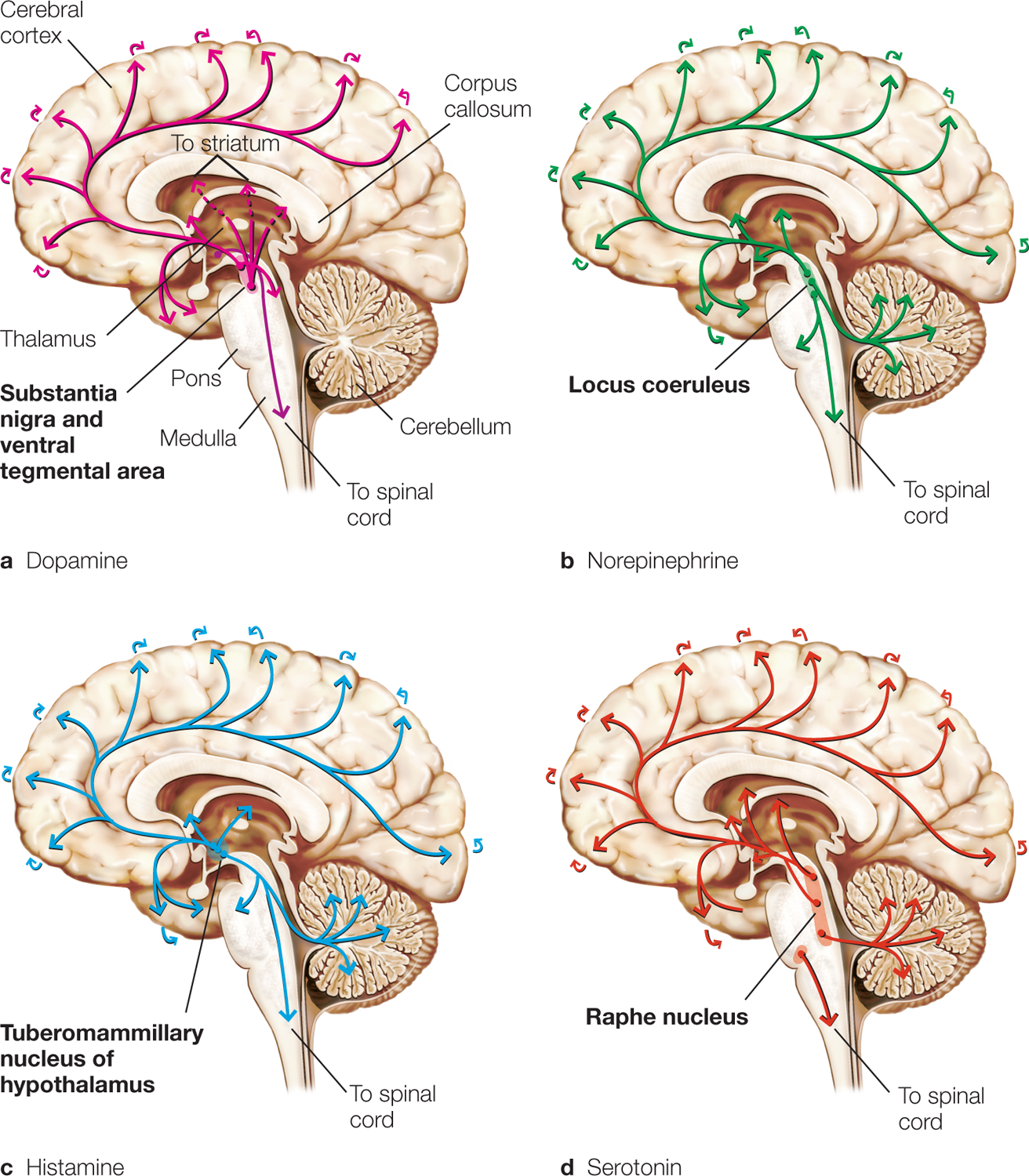
FIGURE 2.15 Major projection pathways of the biogenic amine neurotransmitter systems in the human brain.
Shown are the projections of the dopamine (a), norepinephrine (b), histamine (c), and serotonin (d) systems using the neurotransmitters. The views are midsagittal cuts through the human brain, showing the medial surface of the right hemisphere; the frontal pole is at left. In each image, the primary source of the biogenic amine is in bold type.
Functional classification of neurotransmitters. As mentioned earlier, the effect of a neurotransmitter on the postsynaptic neuron is determined by the postsynaptic receptor’s properties rather than by the transmitter itself. A particular neurotransmitter may have more than one type of postsynaptic receptor to which it binds, mediating different responses. Thus, the same neurotransmitter released from the same presynaptic neuron onto two different postsynaptic cells might cause one to increase firing and the other to decrease firing, depending on the receptors to which the transmitter binds.
The effects also depend on the concentration of the transmitter; the type, number, and density of the receptors; whether, when, and which co-neurotransmitters are also released; and the long-range connections of the neuron. For example, if different co-transmitters are released—one binding to a directly gated receptor with fast signaling and the other to a GPCR with slower signaling—they may produce opposite actions, and their overall combined effect may produce many possible outcomes, even complementary effects. Nevertheless, neurotransmitters can be classified not only biochemically, but also by the typical effect that they induce in the postsynaptic neuron.
Neurotransmitters that usually have an excitatory effect include ACh, the catecholamines, glutamate, histamine, serotonin, and some of the neuropeptides. Neurotransmitters that are typically inhibitory include GABA, glycine, and some of the neuropeptides. Some neurotransmitters act directly to excite or inhibit a postsynaptic neuron, but other neurotransmitters act only in concert with other factors. These are sometimes referred to as conditional neurotransmitters because their action is conditioned on the presence of another transmitter in the synaptic cleft or activity in the neural circuit. These types of mechanisms permit the nervous system to achieve complex modulations of information processing by modulating neurotransmission.
Some common neurotransmitters and their functions. The primary players in the balancing act between excitation and inhibition are glutamate and GABA. Glutamate is released by the pyramidal cells of the cortex, the most common cortical neurons. As a result, glutamate is the most prevalent neurotransmitter and is found in most of the fast excitatory synapses in the brain and spinal cord. A few different types of receptors bind glutamate, and some of these are found in modifiable synapses (i.e., ones that can change in strength) involved in learning and memory. Too much glutamate (excitation) can be toxic and cause cell death and has been implicated in stroke, epilepsy, and neurodegenerative diseases such as Alzheimer’s and Parkinson’s.
GABA is the second most prevalent neurotransmitter and is synthesized from glutamate. It is found in most of the fast inhibitory synapses across the brain. As with glutamate, there is more than one type of GABA receptor, but the most common one opens Cl- channels to allow an influx of negatively charged ions into the cell, negatively shifting (hyperpolarizing) the membrane potential and, in essence, inhibiting the neuron by making it much less likely to fire. GABA’s role in information processing is varied and complex and is actively being researched.
Over the past few years, several populations of neurons that researchers believed released only glutamate, acetylcholine, dopamine, or histamine have also been found to release GABA (reviewed in Tritsch et al., 2016). For example, although glutamate and GABA have opposing functions, their co-release has recently been documented from individual CNS axons (in the ventral tegmental area and entopeduncular nucleus). Defects in the GABA system may be local or affect the entire CNS. Decreased levels of GABA (decreased inhibition) can result in seizures, as well as increases in emotional reactivity, heart rate, blood pressure, food and water intake, sweating, insulin secretion, gastric acid, and colonic motility. Too much GABA can lead to coma.
Acetylcholine (ACh) is present in the synapses between neurons and between neurons and muscles (neuromuscular junctions), where it has an excitatory effect and activates muscles. In the brain, ACh acts as a neurotransmitter and a neuromodulator and supports cognitive function. It binds to two main types of receptors, nicotinic and muscarinic, that have different properties and mechanisms. There are muscle-type and neuronal-type nicotinic ACh receptors; the latter type are located in the autonomic ganglia of the sympathetic and parasympathetic nervous systems (discussed later in this chapter). Nicotine also binds to these receptors and imitates the actions of ACh—hence their name. Muscarinic ACh receptors are found in both the central nervous system and in the heart, lungs, upper gastrointestinal tract, and sweat glands. Some ACh receptors have inhibitory effects; others have excitatory effects. In the CNS, the drug nicotine binds to nicotinic acetylcholine receptors to increase arousal, sustain attention, enhance learning and memory, and increase REM sleep.
Many plants and animals produce toxins and venoms that affect ACh levels. For example, botulinum toxin suppresses the release of ACh at the neuromuscular junction, causing flaccid paralysis. Because of this property, Botox, a commercial product containing minuscule amounts of botulinum toxin, is used to relax the muscles in multiple disorders characterized by overactive muscle activity, such as post-stroke spasticity. It is also used for cosmetic purposes, because a small injection into subcutaneous muscles results in reduced wrinkling of the overlying skin in areas such as around the eyes. The plant toxin curare has a different mechanism. It binds to the nicotinic ACh receptor, also decreasing ACh levels, and causing muscle weakness and, in sufficient doses, flaccid paralysis of the diaphragm and death. Some toxins and venoms inhibit the action of the ACh breakdown enzyme acetylcholinesterase (AChE), leading to excess ACh at the neuromuscular junction and resulting in continuous activation of muscles, causing rigid paralysis. In this case, death can be caused by hyperactivity and rigidity of the muscles needed for breathing.
The primary sites of dopamine production are the adrenal glands and a few small areas of the brain. Brain areas with significant dopaminergic innervation include the striatum, substantia nigra, and hypothalamus. So far, five different types of dopamine receptors have been identified (with hints of two more), labeled from D1 to D5, all of which are G protein–coupled receptors, exerting their effects on postsynaptic neurons via the second-messenger mechanism. There are several dopaminergic pathways, each sprouting from one of the small brain areas where it is produced and each involved in particular functions, including cognitive and motor control, motivation, arousal, reinforcement, and reward, among others. Parkinson’s disease, schizophrenia, attention deficit hyperactivity disorder, and addiction are associated with deficits in dopamine systems.
Serotonin in the brain is released largely by the neurons of the raphe nuclei, located in the brainstem. Axons from the raphe nuclei neurons extend to most parts of the central nervous system, forming a neurotransmitter system. Serotonin receptors—both ligand-gated ion channels and GPCRs—are found on the cell membrane of neurons and other cell types that mediate both excitatory and inhibitory neurotransmission. Serotonergic pathways are involved in the regulation of mood, temperature, appetite, behavior, muscle contraction, sleep, and the cardiovascular and endocrine systems. Serotonin also has effects on learning and memory. Drugs such as the selective serotonin reuptake inhibitors (SSRIs), used to treat clinical depression, act on the raphe nuclei and their targets in the brain.
Norepinephrine (NE), also known as noradrenaline, is the sympathetic nervous system’s go-to neurotransmitter. It is produced and used by neurons with cell bodies in the locus coeruleus (LC), an area of the brain involved with physiological responses to stress and located in one of the brainstem’s structures, the pons. These neurons have extensive projections to the cortex, cerebellum, and spinal cord. Activity in the LC is low during sleep, runs at a baseline level during awake periods, steps up the action when presented with an attention-drawing stimulus, and strongly activates when potential danger is sensed.
Outside the brain, NE is released by the adrenal glands. There are two types of receptors for NE: alpha (α1 and α2) and beta (β1, β2, and β3), both of which are GPCRs. Alpha-2 receptors tend to have inhibitory effects, while alpha-1 and the beta receptors tend to have excitatory effects. NE mediates the fight-or-flight response. Its general effect is to prepare the body and its organs for action. It increases arousal, alertness, and vigilance; focuses attention; and enhances memory formation. Along with these effects come increased anxiety and restlessness. The body responds to NE with increased heart rate, blood pressure, and blood flow to skeletal muscles, and decreased blood flow to the gastrointestinal system. It also increases the availability of stored glucose for energy.
Neurosteroids are steroids synthesized in the brain. Only in the last 40 years have researchers found evidence that the brain can synthesize steroids. There are many different neurosteroids, some inhibitory and some excitatory; they can modulate the binding of various neurotransmitters on both directly and indirectly gated receptors and also directly activate GPCRs (reviewed in Do Rego et al., 2009). Neurosteroids are involved with the control of various neurobiological processes, including cognition, stress, anxiety, depression, aggressiveness, body temperature, blood pressure, locomotion, feeding behavior, and sexual behavior.
For example, estradiol is a hormone derived from cholesterol (like other steroid hormones) and is produced primarily in the ovaries of women and the testes of men. The brain, however, also has the molecules and enzymes necessary for the conversion of cholesterol into steroids such as estradiol (as well as progesterone and testosterone), and it has specific receptors for each of these neurosteroids and for peripherally produced steroid hormones. Estradiol is a neuroprotective factor, and recent discoveries indicate that neural estrogen receptors coordinate multiple signaling mechanisms that protect the brain from neurodegenerative diseases, affective disorders, and cognitive decline (reviewed in Arevalo et al., 2015).
Inactivation of neurotransmitters after release. Following the release of neurotransmitter into the synaptic cleft and its binding with the postsynaptic membrane receptors, the remaining transmitter must be removed to prevent further excitatory or inhibitory signal transduction. This removal can be accomplished by active reuptake of the substance back into the presynaptic terminal, by enzymatic breakdown of the transmitter in the synaptic cleft, or merely by diffusion of the neurotransmitter away from the region of the synapse or site of action (e.g., in the case of hormones that act on target cells distant from the synaptic terminals).
Neurotransmitters that are removed from the synaptic cleft by reuptake mechanisms include the biogenic amines (dopamine, norepinephrine, epinephrine, histamine, and serotonin). The reuptake mechanism is mediated by active transporters, which are transmembrane proteins that pump the neurotransmitter back across the presynaptic membrane.
An example of a neurotransmitter that is eliminated from the synaptic cleft by enzymatic action is ACh. The enzyme AChE, located in the synaptic cleft, breaks down ACh after it has acted on the postsynaptic membrane. In fact, special AChE stains (chemicals that bind to AChE) can be used to label AChE on muscle cells, thus revealing where motor neurons innervate the muscle.
To monitor the level of neurotransmitter in the synaptic cleft, presynaptic neurons have autoreceptors. These autoreceptors are located on the presynaptic terminal and bind with the released neurotransmitter, enabling the presynaptic neuron to regulate the synthesis and release of the transmitter.
Electrical Transmission
Some neurons communicate via electrical synapses, which are very different from chemical synapses. In electrical synapses, no synaptic cleft separates the neurons. Instead, the neuronal membranes touch at specializations called gap junctions, and the cytoplasms of the two neurons are essentially continuous. These gap junction channels create pores connecting the cytoplasms of the two neurons (Figure 2.16). As a result, the two neurons are isopotential (i.e., they have the same electrical potential), meaning that electrical changes in one are reflected instantaneously in the other. Following the principles of electrotonic conduction, however, the passive currents that flow between the neurons when one of them is depolarized (or hyperpolarized) decrease and are therefore smaller in the postsynaptic neuron than in the presynaptic neuron. Under most circumstances, the communication is bidirectional; however, so-called rectifying synapses limit current flow in one direction, as is typical in chemical synapses.
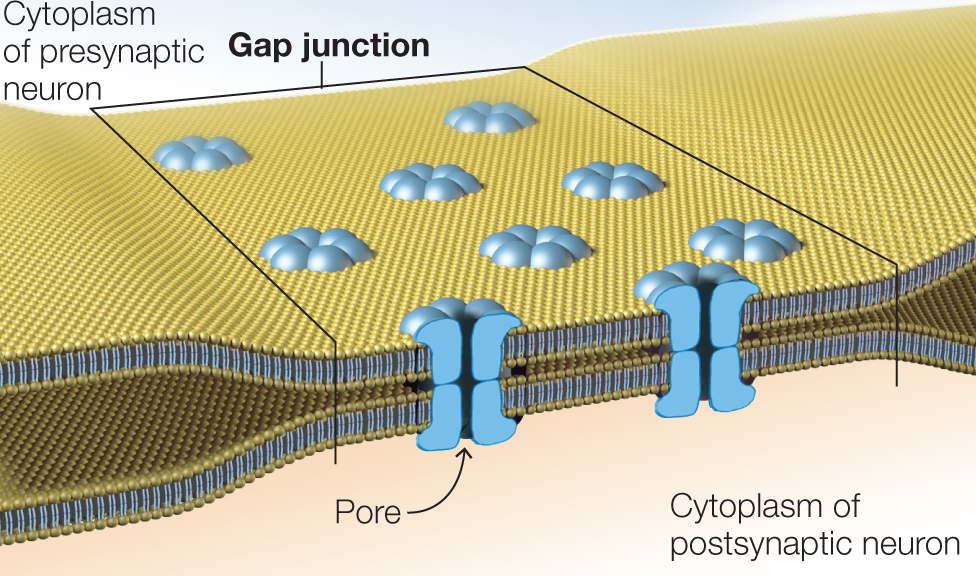
FIGURE 2.16 Electrical synapse between two neurons.
Electrical synapses are formed by gap junctions, places where multiple transmembrane proteins in the pre- and postsynaptic neurons connect to create pathways that connect the cytoplasms of the two neurons.
Electrical synapses are useful when information must be conducted rapidly, such as in the escape reflex of some invertebrates. Groups of neurons with these synapses can activate muscles quickly to get the animal out of harm’s way. For example, the well-known tail flip reflex of crayfishes involves powerful rectifying electrical synapses. Electrical synapses are also useful when groups of neurons should operate synchronously, as with some hypothalamic neurosecretory neurons. Electrical synapses also have some limitations: They are much less plastic than chemical synapses, and they cannot amplify a signal (whereas an action potential that triggers a chemical synapse could cause a large release of neurotransmitter, thus amplifying the signal).
Take-Home Messages
- Synapses are the locations where one neuron can transfer information to another neuron or a specialized nonneuronal cell. They are found on dendrites and at axon terminals but can also be found on the neuronal cell body.
- Chemical transmission results in the release of neurotransmitters from the presynaptic neuron and the binding of those neurotransmitters on the postsynaptic neuron, which in turn causes excitatory or inhibitory postsynaptic potentials (EPSPs or IPSPs), depending on the properties of the postsynaptic receptor.
- Neurotransmitters must be removed from the receptor after binding. This removal can be accomplished by active reuptake back into the presynaptic terminal, enzymatic breakdown of the transmitter in the synaptic cleft, or diffusion of the neurotransmitter away from the region of the synapse.
- Electrical synapses are different from chemical synapses because they operate by passing current directly from one neuron (presynaptic) to another neuron (postsynaptic) via specialized channels in gap junctions that connect the cytoplasm of one cell directly to the other.
- synaptic cleft
The gap between neurons at synapses.
- vesicle
A small intracellular organelle, located in the presynaptic terminals at synapses, that contains neurotransmitter.
- neurotransmitter
A chemical substance that transmits the signal between neurons at chemical synapses.