2.7 Development of the Nervous System
Thus far, we have been discussing the neuroanatomy of the developed adult brain. In humans and many other species, the fetal brain is well developed and shows cortical layers, neuronal connectivity, and myelination; in short, the fetal brain is already extremely complex, although by no means completely developed. This section provides an introduction to development, with special attention to development of the cerebral cortex.
Overview of Early Development
Fertilization of the egg is followed by a series of events that lead to the formation of a multicellular blastula, which has already begun to specialize. The blastula contains three main cell lines that, after a few days, form three layers: the ectoderm (outer layer) that will form the nervous system and the outer skin, lens of the eye, inner ear, and hair; the mesoderm (middle layer) that will form the skeletal system and voluntary muscle; and the endoderm (inner layer) that will form the gut and digestive organs. The early processes that go into forming the nervous system are called neurulation (Figure 2.45). During this stage, the ectodermal cells on the dorsal surface form the neural plate.
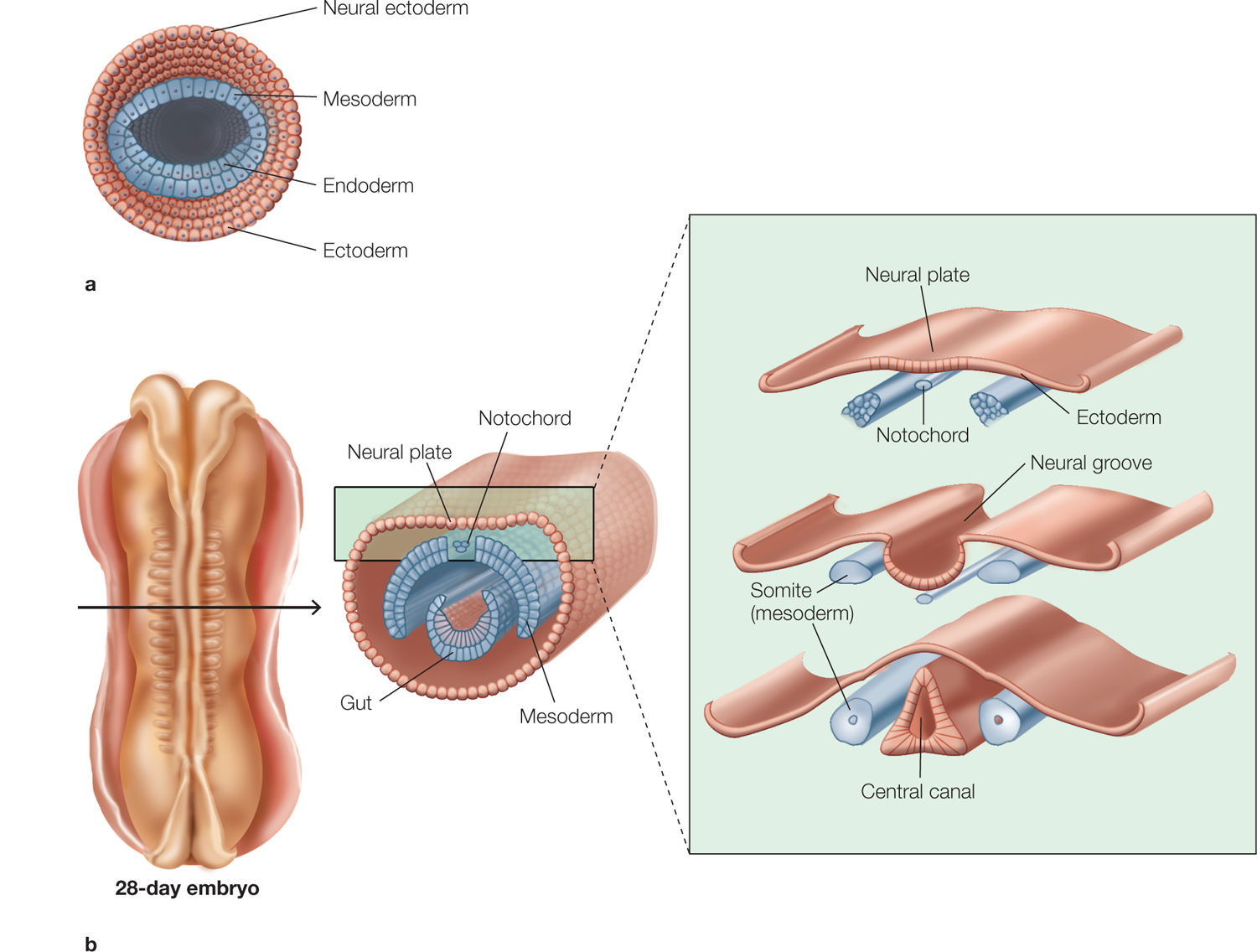
FIGURE 2.45 Development of the vertebrate nervous system.
Cross sections through the blastula and embryo at various developmental stages during the first 21 days of life. (a) Early in embryogenesis, the multicellular blastula contains cells destined to form various body tissues. (b) Migration and specialization of different cell lines leads to formation of the primitive nervous system around the neural groove and, after it fuses, the neural tube (not shown) on the dorsal surface of the embryo. The brain is located at the anterior end of the embryo and is not shown in these more posterior sections, which are taken at the level of the spinal cord.
As the nervous system continues to develop, the cells at the lateral borders of the neural plate push upward. (Imagine joining the long sides of a rectangular piece of dough to form a tube.) This movement causes the more central cells of the neural plate to invaginate, or dip inward, to form the neural groove. As the groove deepens, the cells pushing up at the border of the neural fold region eventually meet and fuse, forming the neural tube that runs anteriorly and posteriorly along the embryo. The adjacent nonneural ectoderm then reunites to seal the neural tube within an ectodermal covering that surrounds the embryo.
At both ends of the neural tube are openings (the anterior and the posterior neuropores) that close on about the 23rd to 26th day of gestation. When the anterior neuropore is sealed, this cavity forms the primitive brain, consisting of three spaces, or ventricles. If the neuropores do not close correctly, neural tube defects such as anencephaly (absence of a major portion of the brain and skull) or spina bifida (incomplete formation of some of the vertebrae) may result. From this stage on, the brain’s gross features are formed by growth and flexion (bending) of the neural tube’s anterior portions (Figure 2.46). The result is a cerebral cortex that envelops the subcortical and brainstem structures. The final three-dimensional relations of the brain’s structures are the product of continued cortical enlargement and folding. The posterior portion of the neural tube differentiates into a series of repeated segments that form the spinal cord.
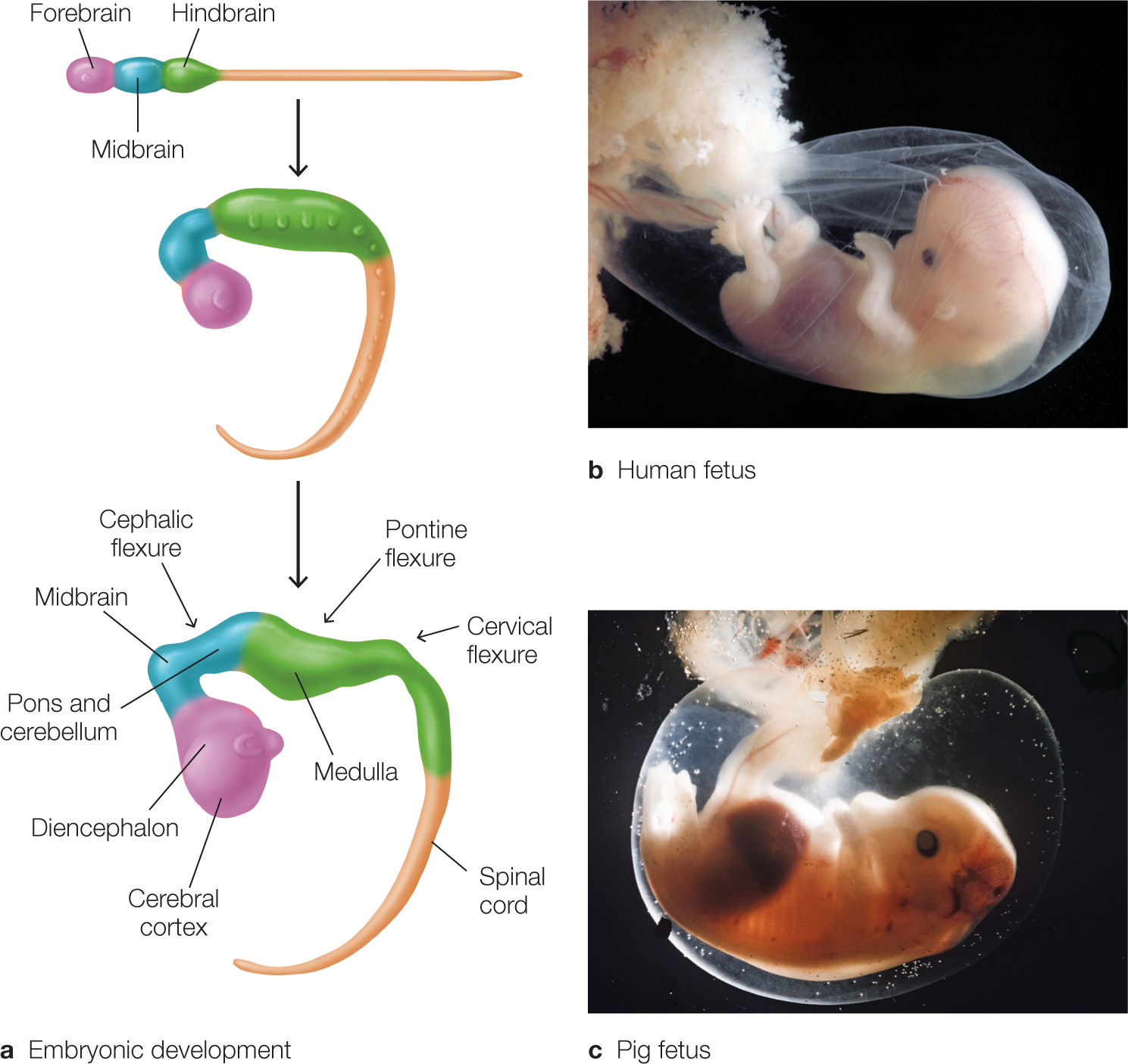
FIGURE 2.46 Early stages of embryonic development in mammals.
(a) Developing embryo. The embryo goes through a series of folds, or flexures, during development. These alterations in the gross structure of the nervous system give rise to the compact organization of the adult brain and brainstem in which the cerebral cortex overlies the diencephalon and midbrain within the human skull. There is significant similarity between the gross features of the developing fetuses of mammals, as comparison of a human fetus (b) and a pig fetus (c) shows.
In primates, almost the full complement of neurons is generated prenatally during the middle third of gestation. The entire adult pattern of gross and cellular neural anatomical features is present at birth, and very few neurons are generated after birth (but see the end of this section, “Birth of New Neurons Throughout Life”). Although axonal myelination continues for some period postnatally (e.g., until adulthood in the human frontal lobe), the newborn has a well-developed cortex that includes the cortical layers and areas characterized in adults. For instance, BA17 (the primary visual cortex) can be distinguished from the motor cortex by cytoarchitectonic analysis of its neuronal makeup at the time of birth.
Neural Proliferation and Migration of Cortical Cells The neurons that form the brain arise from a layer of precursor cells in proliferative zones located adjacent to the ventricles of the developing brain. The cortical neurons arise from the subventricular zone, and those that form other parts of the brain arise from precursor cells in the ventricular zone. Figure 2.47 shows a cross section through the cortex and the precursor cell layers at various times during gestation. The cells that form the cortex will be the focus of this discussion. The precursorcells are undifferentiated cells from which all cortical cells (including neuronal subtypes and glial cells) arise through cell division and differentiation. For the first 5 to 6 weeks of gestation, the cells in the subventricular zone divide in a symmetrical fashion. The result is exponential growth in the number of precursor cells.
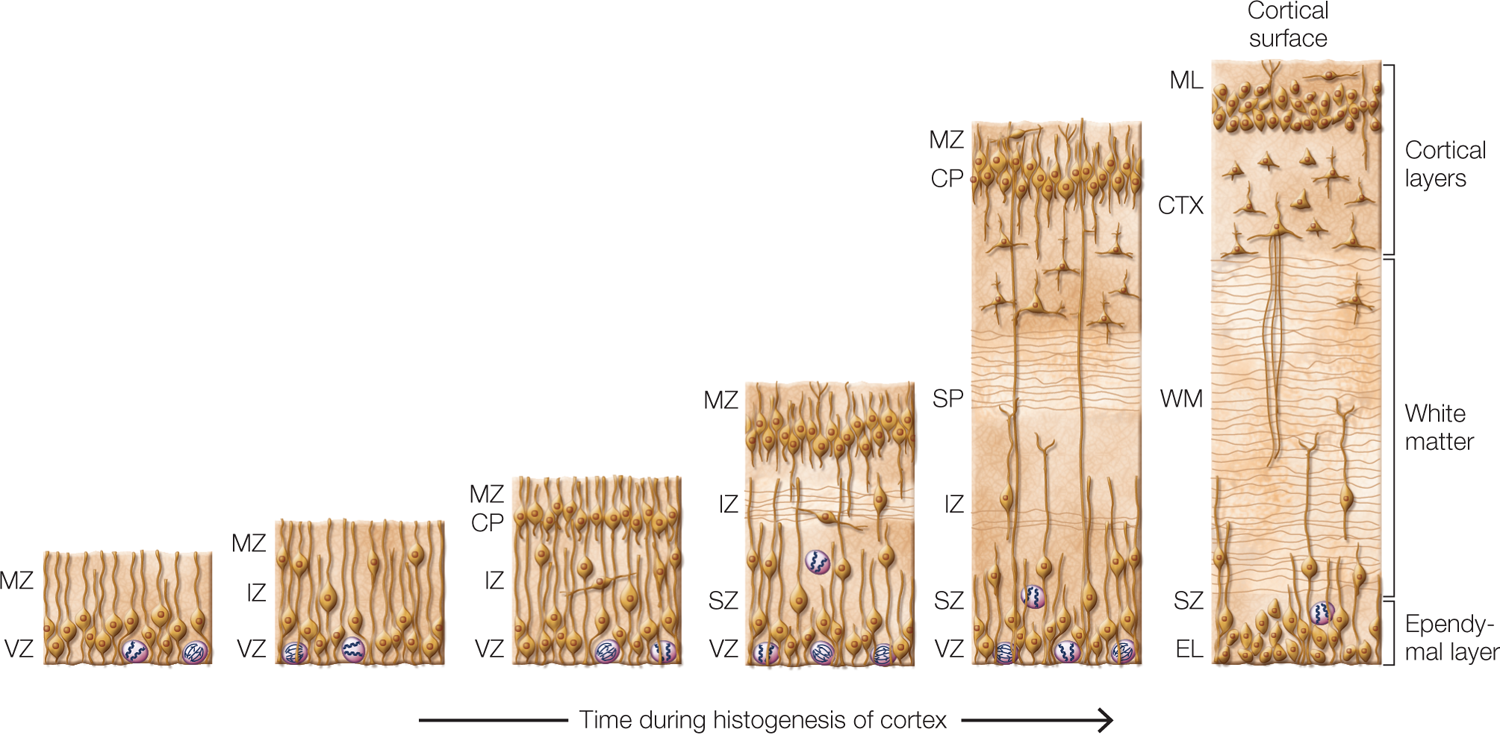
FIGURE 2.47 Histogenesis of the cerebral cortex.
Cross-sectional views of developing cerebral cortex from early (left) through late (right) times during histogenesis. The mammalian cortex develops from the inside out as cells in the ventricular zone (VZ) divide, and some of the cells migrate to the appropriate layer in the cortex. Radial glial cells form a superhighway along which the migrating cells travel en route to the cortex. CP = cortical plate; CTX = cortex; EL = ependymal layer; IZ = intermediate zone; ML = molecular layer; MZ = marginal zone; SP = subplate; SZ = subventricular zone; WM = white matter.
At the end of 6 weeks, when there is a stockpile of precursor cells, asymmetrical division begins. After every cell division, one of the two cells formed becomes a migratory cell destined to be part of another layer; the other cell remains in the subventricular zone, where it continues to divide asymmetrically. Later in gestation, the proportion of migratory cells increases until a laminar (i.e., layered) cortex made up of the migratory cells is formed. This cortex has a foundational epithelial layer that becomes the cell lining of the ventricles and is known as the ependymal cell layer.
The migratory cells travel outward from the subventricular zone by moving along peculiar cells known as radial glial cells, which stretch from the subventricular zone to the surface of the developing cortex. The work of radial glial cells does not end with development. These cells are transformed into astrocytes in the adult brain, helping to form part of the blood–brain barrier.
As the first migrating neurons approach the surface of the developing cortex—a point known as the cortical plate—they stop short of the surface. Neurons that migrate later pass beyond the termination point of the initial neurons and end up in more superficial positions—positions nearer the outer cortical surface. Thus, it is said that the cortex is built from the inside out, because the first neurons to migrate lie in the deepest cortical layers, whereas the last to migrate move farthest out toward the cortical surface.
Neuronal Determination and Differentiation The cortex is made up of many different types of neurons organized in a laminar fashion. Layer IV, for example, contains large pyramidal cells, layer III is populated primarily by stellate cells, and so on. You may be wondering how that population of virtually identical precursor cells gives rise to the variety of neurons and glial cells in the adult cortex. What determines the type of neuron that a migrating cell is fated to become?
The answer lies in the timing of neurogenesis. Experimental manipulation of developing cells has shown that the differentiated cell type is not hardwired into the code of each developing neuron. Neurons that are experimentally prevented from migrating, by being exposed to high-energy X-rays, eventually form cell types and patterns of connectivity that would be expected from neurons created at the same gestational stage. Even though the thwarted neurons might remain in the ventricular zone, they display interconnections with other neurons that would be normal if they had migrated to the cortical layers normally.
The timeline of cortical neurogenesis differs across cortical cytoarchitectonic areas, but the inside-out pattern is the same for all cortical areas. Because the timeline of cortical neurogenesis determines the ultimate pattern of cortical lamination, anything that affects the genesis of cortical neurons will lead to an ill-constructed cortex. A good example of how neuronal migration can be disrupted in humans is fetal alcohol syndrome. In cases of chronic maternal alcohol abuse, neuronal migration is severely disrupted and results in a disordered cortex, leading to a plethora of cognitive, emotional, and physical disabilities.
The Baby Brain: Ready to Rock ’n’ Roll?
A host of behavioral changes takes place during the first months and years of life. What accompanying neurobiological changes enable these developments? Even if we assume that neuronal proliferation continues, we know that at birth, the human brain has a fairly full complement of neurons, and these are organized to form a human nervous system that is normal, even if not complete in all details. Which details are incomplete, and what is known about the time course of brain maturation?
Although the brain nearly quadruples in size from birth to adulthood, the change is not due to an increase in neuron number. A substantial amount of that growth comes from synaptogenesis (the formation of synapses) and the growth of dendritic trees. Synapses in the brain begin to form long before birth—before week 27 in humans (counting from conception)—but they do not reach peak density until after birth, during the first 15 months of life. Synaptogenesis is more pronounced early in the deeper cortical layers and occurs later in more superficial layers, following the pattern of neurogenesis described earlier.
At roughly the same time that synaptogenesis is occurring, neurons of the brain are increasing the size of their dendritic arborizations, extending their axons, and undergoing myelination. Synaptogenesis is followed by synapse elimination (sometimes called pruning), which continues for more than a decade. Synapse elimination is a means by which the nervous system fine-tunes neuronal connectivity, presumably eliminating the interconnections between neurons that are redundant, unused, or do not remain functional.
Use it or lose it! The person you become is shaped by the growth and elimination of synapses, which in turn are shaped by the world you’re exposed to and the experiences you have. A specific example of synaptic pruning comes from primary visual cortex (BA17): Initially, there is overlap between the projections of the two eyes onto neurons in BA17. After synapse elimination, the cortical inputs from the two eyes within BA17 are nearly completely segregated. The axon terminals relaying information from each eye form a series of equally spaced patches (called ocular dominance columns), and each patch receives inputs from predominantly one eye.
One of the central hypotheses about the process of human synaptogenesis and synapse elimination is that the time course of these events differs in different cortical regions. The data suggest that in humans, synaptogenesis and synapse elimination peak earlier in sensory (and motor) cortex than in association cortex. By contrast, in the brain development of other primates, synaptogenesis and pruning appear to occur at the same rates across different cortical regions. Differences in methodology, however, must be resolved before these interspecies variations will be wholly accepted. Nonetheless, compelling evidence suggests that different regions of the human brain reach maturity at different times.
The increase in brain volume that occurs postnatally is also a result of both myelination and the proliferation of glial cells. White matter volume increases linearly with age across cortical regions (Giedd et al., 1999). In contrast, gray matter volume increases nonlinearly, showing a preadolescent increase followed by a postadolescent decrease. In addition, the time courses of gray matter increase and decrease are not the same across different cortical regions. In general, these data support the idea that postnatal developmental changes in the human cerebral cortex may not occur with the same time course across all cortical regions (see also Shaw et al., 2006).
Birth of New Neurons Throughout Life
One principle about the human brain that once dominated in the neuroscience community was the idea that the adult brain produces no new neurons. This view was held despite a variety of claims of neurogenesis in the brain in histological studies dating as far back as the time of Ramón y Cajal. In the last couple of decades, however, studies using an array of modern neuroanatomical techniques have challenged this belief.
Neurogenesis in adult mammals has now been well established in two brain regions: the hippocampus and the olfactory bulb. Neurogenesis in the hippocampus is particularly noteworthy because it plays a key role in learning and memory. In rodents, studies have shown that stem cells in a region of the hippocampus known as the dentate gyrus produce new neurons in the adult, and these can migrate into regions of the hippocampus where similar neurons are already functioning. It is important to know that these new neurons can form dendrites and send out axons along pathways expected of neurons in this region of the hippocampus, and they can also show signs of normal synaptic activity. These findings are particularly interesting because the number of new neurons correlates positively with learning or enriched experience (more social contact or challenges in the physical environment) and negatively with stress (e.g., living in an overcrowded environment). Moreover, the number of newborn neurons is related to hippocampal-dependent memory (Shors, 2004).
Other investigators have found that these new neurons become integrated into functional networks of neurons and participate in behavioral and cognitive functions in the same way that those generated during development do (Ramirez-Amaya et al., 2006). Future work will be required to establish whether adult neurogenesis occurs more broadly in the mammalian brain or is restricted to the olfactory bulb and hippocampus.
What about the adult human brain? Does neurogenesis also occur in mature humans? In a fascinating line of research, a team of scientists from California and Sweden (Eriksson et al., 1998) explored this question in a group of terminally ill cancer patients. As part of a diagnostic procedure related to their treatment, the patients were given BrdU, a synthetic form of thymidine used as a label to identify neurogenesis. The purpose was to assess the extent to which the tumors in the cancer patients were proliferating; tumor cells that were dividing would also take up BrdU, and this label could be used to quantify the progress of the disease.
Neurons undergoing mitotic division during neurogenesis in these patients also took up BrdU, which could be observed in postmortem histological examinations of their brains. The postmortem tissue was immunostained to identify neuron-specific cell surface markers. The scientists found cells labeled with BrdU in the subventricular zone of the caudate nucleus and in the granular cell layer of the dentate gyrus of the hippocampus (Figure 2.48). By staining the tissue to identify neuronal markers, the researchers showed that the BrdU-labeled cells were neurons (Figure 2.49). These findings demonstrate that new neurons are produced in the adult human brain, and that our brains renew themselves throughout life to an extent not previously thought possible.
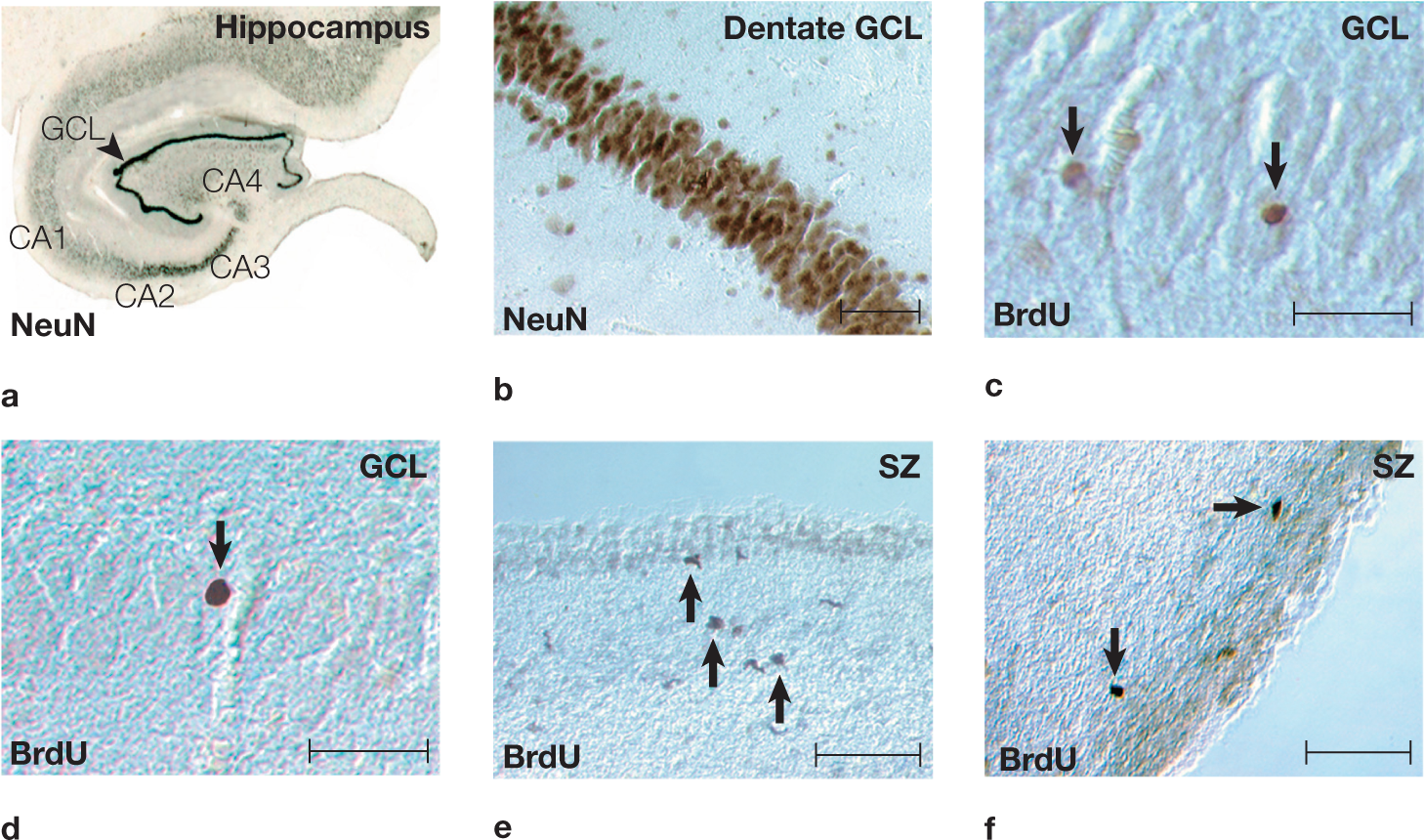
FIGURE 2.48 Newly born neurons in adult human brain tissue.
(a) The hippocampus of the adult human brain, stained for a neuronal marker (NeuN). (b) The dentate gyrus granule cell layer (GCL) in a NeuN-stained section. (c) BrdU-labeled nuclei (arrows) in the granule cell layer of the dentate gyrus. (d) BrdU-labeled cells (arrow) in the granule cell layer of the dentate gyrus. (e) BrdU-stained cells (arrows) adjacent to the ependymal lining in the subventricular zone (SZ) of the human caudate nucleus. These neurons have elongated nuclei resembling the migrating cells that typically are found in the rat subventricular zone. (f) BrdU-stained cells (arrows) with round to elongated nuclei in the subventricular zone of the human caudate nucleus. Scale bars throughout = 50 µm.
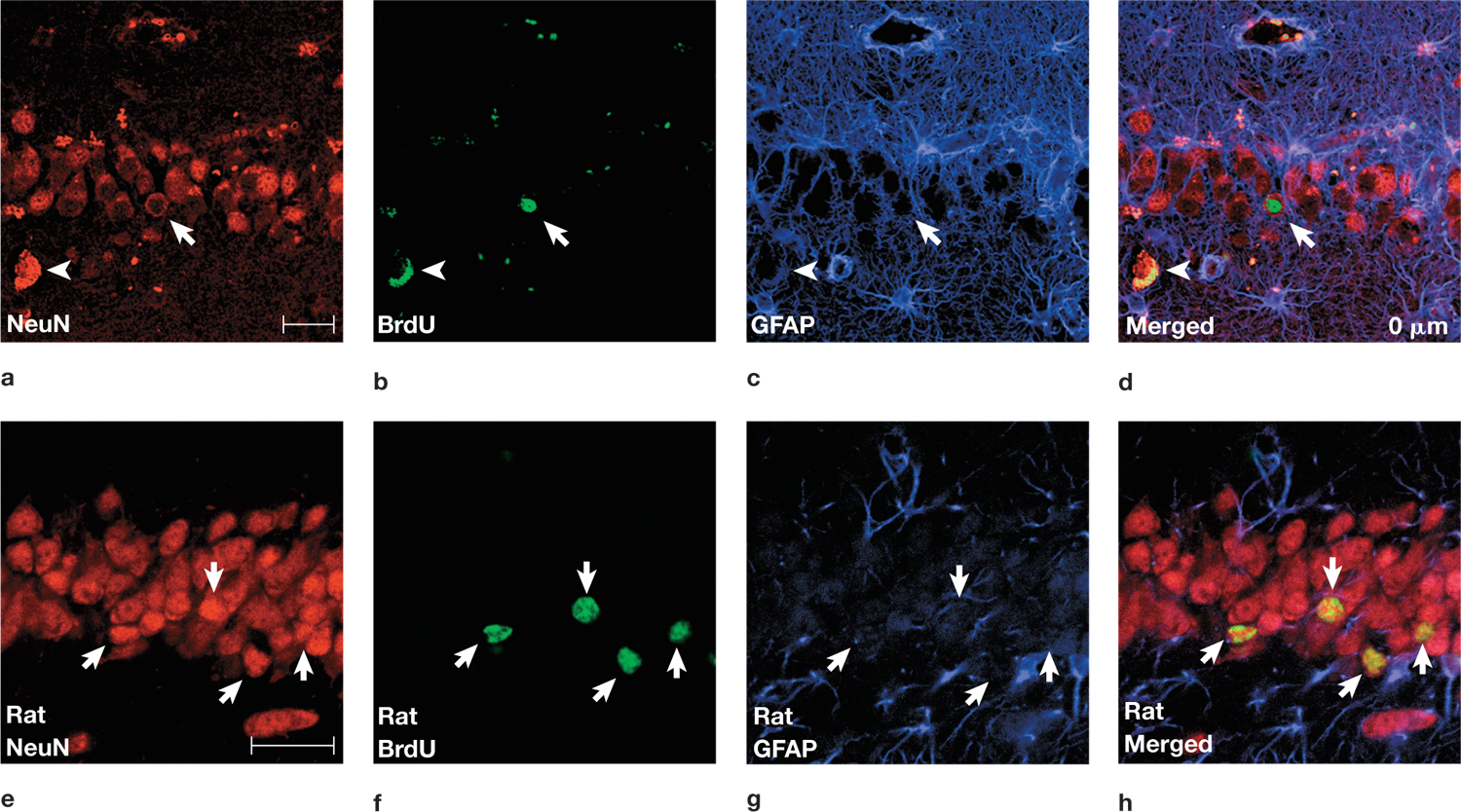
FIGURE 2.49 The birth of new neurons in the dentate gyrus of the adult human (a–d) compared to those in the adult rat (e–h).
New neurons show simultaneous labeling for different stains. (a) A neuron is labeled for NeuN, a neuronal marker. (b) The same cell is labeled with BrdU, indicating that it is newly born (full arrow). (Note that the lone arrowheads in (a) through (d) are pointing to neurons that are fluorescing red or green, owing to nonspecific staining; i.e., these are not newly born neurons.) (c) This same cell is not stained by glial fibrillary acidic protein (GFAP), indicating that it is not an astrocyte. (d) The three stained sections are merged in this image, which shows that a BrdU-labeled cell could specifically coexpress NeuN without expressing GFAP. Panels (e) through (h) show the similarity of the BrdU-labeled neurons in rat dentate gyrus. Note: The scale bar is equal to 25 µm for both sets of images, so the magnification is greater in (e) through (h) than in (a) through (d).
These exciting results hold great promise for the future of neuroscience. Research is under way to investigate the functionality of new neurons in the adult brain and to determine whether such neuronal growth can be facilitated to ameliorate brain damage or the effects of diseases such as Alzheimer’s.
Take-Home Messages
- Neuronal proliferation is the process of cell division in the developing embryo and fetus. It is responsible for populating the nervous system with neurons.
- Neurons and glial cells are formed from precursor cells. After mitosis, these cells migrate along the radial glial cells to the developing cortex. The type of cell that is made (e.g., a stellate or pyramidal cell) appears to be based on when the cell is born (genesis) rather than when it begins to migrate.
- Synaptogenesis is the birth of new synapses; neurogenesis is the birth of new neurons.
- A belief once strongly held by most neuroscientists was that the adult brain produces no new neurons. We now know that this is not the case; new neurons form throughout life in certain brain regions.
- synaptogenesis
The formation of synaptic connections between neurons in the developing nervous system.
- synapse elimination
Also pruning. The elimination of some synaptic contacts between neurons during development, including postnatally.