G-protein-coupled receptors (GPCRs) form the largest family of cell-surface receptors. There are more than 700 GPCRs in humans, and mice have about 1000 involved in the sense of smell alone. These receptors mediate responses to an enormous diversity of extracellular signal molecules, including hormones, local mediators, and neurotransmitters. The signal molecules that bind GPCRs are as varied in structure as they are in function: they can be proteins, small peptides, or derivatives of amino acids or fatty acids, and for each one of them there is a different receptor or set of receptors. Because GPCRs are involved in such a large variety of cell processes, they are an attractive target for the development of drugs to treat many disorders. More than one-third of all drugs used today work through GPCRs.
Despite the diversity of the signal molecules that bind to them, all GPCRs have a similar structure: each is made of a single polypeptide chain that threads back and forth across the lipid bilayer seven times (Figure 16–14). The GPCR superfamily includes rhodopsin (the light-activated photoreceptor protein in the vertebrate eye), the olfactory (smell) receptors in the vertebrate nose, and the receptors that participate in the mating rituals of single-celled yeasts (see Figure 16–1). Evolutionarily speaking, GPCRs are ancient: even prokaryotes possess structurally similar membrane proteins—such as the bacteriorhodopsin that functions as a light-driven H+ pump (see Figure 11–28). Although they resemble eukaryotic GPCRs, these prokaryotic proteins do not act through G proteins, but are coupled to other signal transduction systems.
More information
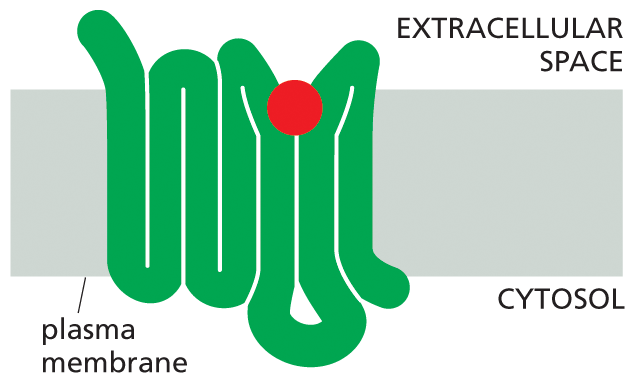
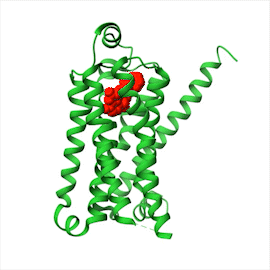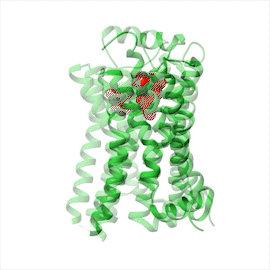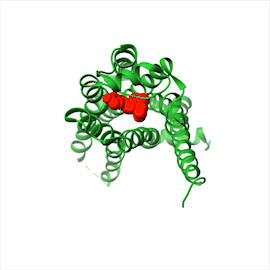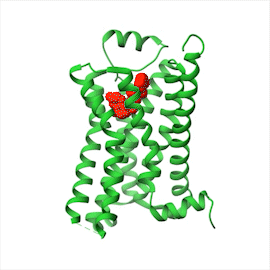
Illustration A shows a folded receptor protein bound to a signal molecule. A folded receptor protein is shown spanning the plasma membrane in a folded structure, and bound to a signal molecule on the extracellular side.
(A)More information
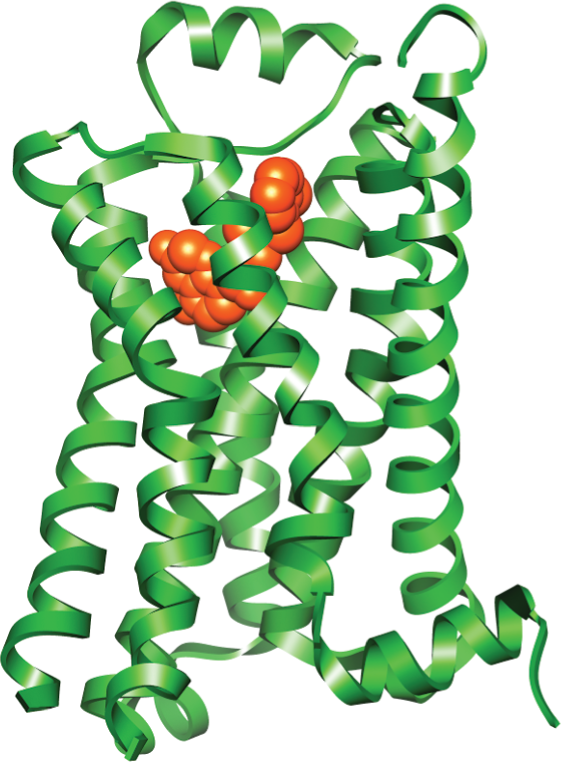
Illustration B shows a protein structure model of a G P C R with a ribbon diagram of epinephrine at the center. G P C R consists of multiple alpha helices, and epinephrine is a small clump in the middle.
(B)Figure 16–14All GPCRs possess a similar structure. The polypeptide chain traverses the membrane as seven α helices. The cytoplasmic portions of the receptor bind to a G protein inside the cell. (A) For receptors that recognize small signal molecules, such as acetylcholine or epinephrine, the ligand (red) usually binds deep within the plane of the membrane to a pocket that is formed by amino acids from several transmembrane segments. Receptors that recognize signal molecules that are proteins usually have a large, extracellular domain that, together with some of the transmembrane segments, binds the protein ligand (not shown). (B) Shown here is the structure of a GPCR that binds to epinephrine (red). Stimulation of this receptor by epinephrine makes the heart beat faster. (Dynamic Figure) The rotating, three-dimensional ribbon structure (below) shows the location of the pocket in which epinephrine (red) binds to a GPCR (green).
We begin this section with a discussion of how G proteins are activated by GPCRs. We then consider how activated G proteins stimulate ion channels and how they regulate membrane-bound enzymes that control the concentrations of small intracellular messenger molecules, including cyclic AMP and Ca2+, which in turn control the activity of important intracellular signaling proteins. We end with a discussion of how light-activated GPCRs in photoreceptors in our eyes enable us to see.
Stimulation of GPCRs Activates G-Protein Subunits
When an extracellular signal molecule binds to a GPCR, the receptor protein undergoes a conformational change that enables it to activate a G protein located on the cytosolic side of the plasma membrane. To explain how this activation leads to the transmission of a signal, we must first consider how G proteins are constructed and how they operate.
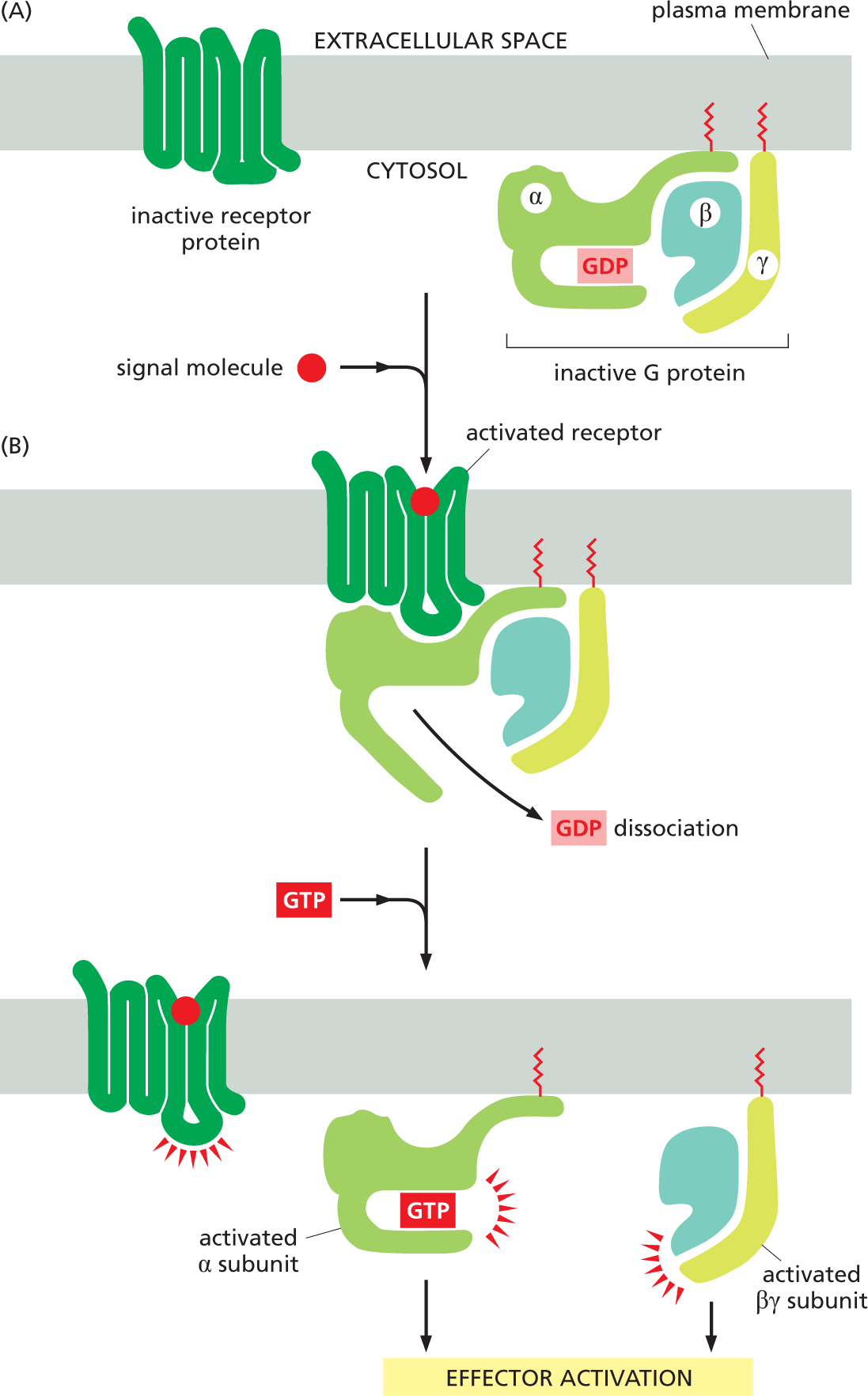
There are several varieties of G proteins. Each is specific for a particular set of receptors and for a particular set of target enzymes or ion channels in the plasma membrane. All of these G proteins, however, have a similar general structure and operate in a similar way. They are composed of three protein subunits—α, β, and γ—two of which are tethered to the plasma membrane by short lipid tails. In the unstimulated state, the α subunit has GDP bound to it, and the G protein is idle (Figure 16–15A). When an extracellular signal molecule binds to its receptor, the altered receptor activates a G protein by causing the α subunit to decrease its affinity for GDP, which is then exchanged for a molecule of GTP. In some cases, this activation breaks up the G-protein subunits, so that the activated α subunit, clutching its GTP, detaches from the βγ complex, which is also activated (Figure 16–15B). The two activated parts of the G protein—the α subunit and the βγ complex—can then each interact directly with target proteins in the plasma membrane, which in turn may relay the signal to other destinations in the cell. The longer these target proteins remain bound to an activated α subunit or a βγ complex, the more prolonged the relayed signal will be.
More information
Two illustrations A and B show the function of G P C R subunits in effector activation. Illustration A depicts an inactive receptor protein bound to the plasma membrane. An inactive G protein with alpha, beta, and gamma units is also attached to the plasma membrane on the cytosolic side. The alpha unit is bound to G D P.
Illustration A leads to illustration B. An extracellular signal molecule binds with a receptor attached to the plasma membrane and activates it. The receptor protein interacts with the inactive G protein. This leads to G D P dissociation from the G protein. In the following step, the receptor on the plasma membrane bound to the signal molecule is activated. G T P binds to the alpha subunit which separates from the activated beta-gamma complex. This leads to effector activation.
Figure 16–15An activated GPCR activates G proteins by encouraging the α subunit to release its GDP and pick up GTP. (A) In the unstimulated state, the receptor and the G protein are both inactive. Although they are shown here as separate entities in the plasma membrane, in some cases they are associated in a preformed complex. (B) Binding of an extracellular signal molecule to the receptor changes the conformation of the receptor, which in turn alters the conformation of the bound G protein. The alteration of the α subunit of the G protein allows it to exchange its GDP for GTP. This exchange triggers an additional conformational change that activates both the α subunit and the βγ complex, which dissociate to interact with their preferred target proteins in the plasma membrane (Movie 16.2). The receptor stays active as long as the external signal molecule is bound to it, and it can therefore activate many molecules of G protein. Note that both the α and γ subunits of the G protein have covalently attached lipid molecules (red) that help anchor the subunits to the plasma membrane.
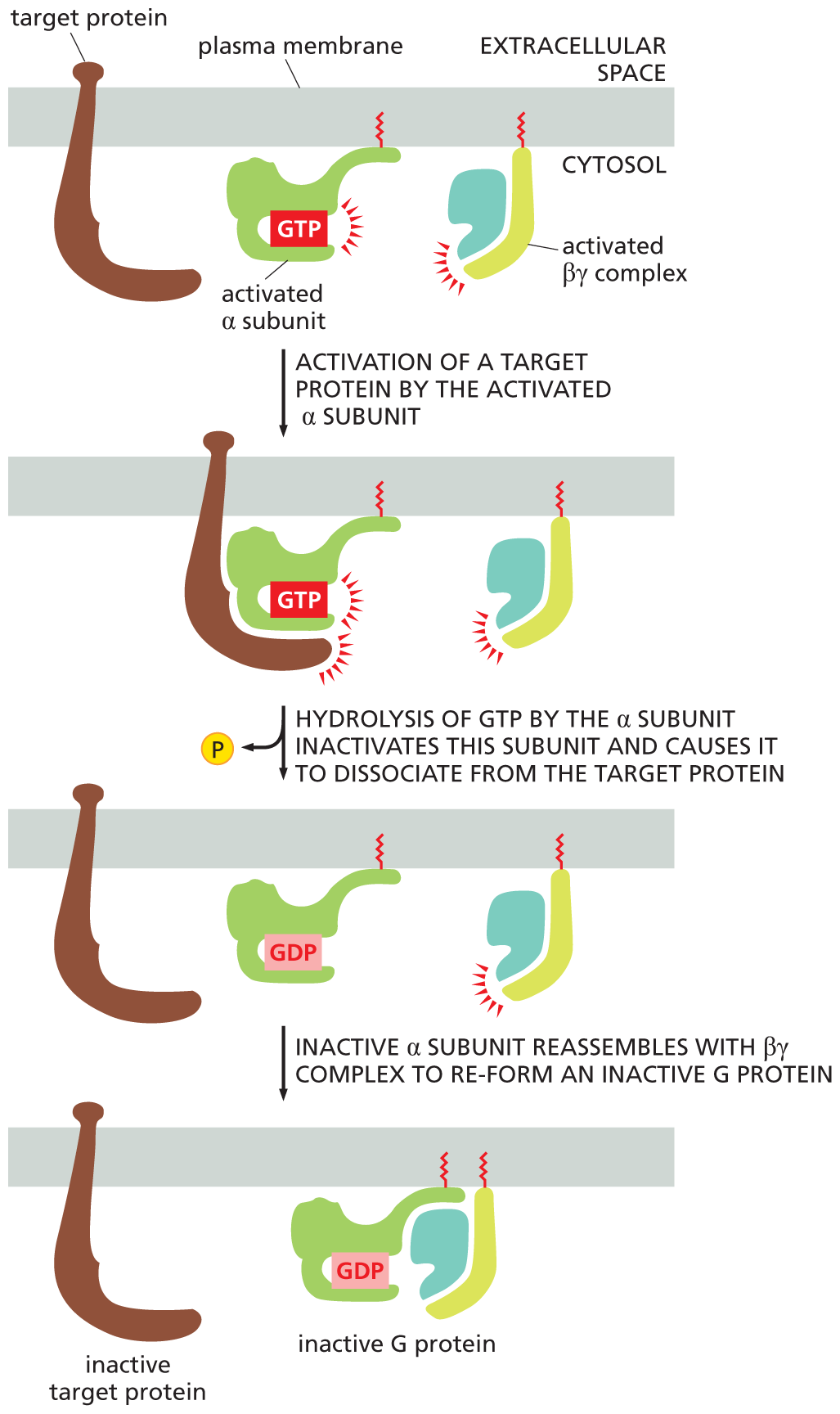
The amount of time that the α subunit and βγ complex remain “switched on”—and hence available to relay signals—also influences how long a response lasts. This timing is controlled by the behavior of the α subunit. The α subunit has an intrinsic GTPase activity, and it eventually hydrolyzes its bound GTP to GDP, returning the whole G protein to its original, inactive conformation (Figure 16–16). GTP hydrolysis and inactivation usually occur within seconds after the G protein has been activated. The inactive G protein is then ready to be reactivated by another activated GPCR.
More information
An illustration depicts the fusion of alpha and beta gamma subunits to form an inactive G protein. A target protein, an activated alpha subunit, and an activated beta-gamma complex are attached to the plasma membrane.
In the first step, activation of a target protein by the activated alpha subunit is shown. The target protein is bound to the activated alpha subunit. An activated beta gamma complex is shown separate from the alpha subunit, and remains separate until the third step.
In the second step, hydrolysis of G T P by the alpha subunit inactivates this subunit and causes it to dissociate from the target protein. The phosphate molecule is released.
In the third step, an inactive alpha subunit reassembles with the beta gamma complex to re-form an inactive G protein. The schematic shows the inactive target protein attached to the plasma membrane separately, and the alpha subunit with G D P bound to the beta gamma units.
Figure 16–16The G protein α subunit switches itself off by hydrolyzing its bound GTP to GDP. When an activated α subunit interacts with its target protein, it activates that target protein for as long as the two remain in contact. (In some cases, this interaction will inactivate the target protein; not shown.) The α subunit then hydrolyzes its bound GTP to GDP—an event that takes place usually within seconds of G-protein activation. The hydrolysis of GTP inactivates the α subunit, which dissociates from its target protein and—if the α subunit had separated from the βγ complex (as shown)—reassociates with a βγ complex to re-form an inactive G protein (see also Movie 16.2). The G protein is now ready to couple to another activated receptor, as in Figure 16–15B. Both the activated α subunit and the activated βγ complex can interact with target proteins in the plasma membrane.
Some Bacterial Toxins Cause Disease by Altering the Activity of G Proteins
QUESTION 16–3
GPCRs activate G proteins by reducing the strength of GDP binding to the G protein. This results in rapid dissociation of bound GDP, which is then replaced by GTP, because GTP is present in the cytosol in much higher concentrations than GDP. What consequences would result from a mutation in the α subunit of a G protein that caused its affinity for GDP to be reduced without significantly changing its affinity for GTP? Compare the effects of this mutation with the effects of cholera toxin.
G proteins offer a striking example of the importance of being able to shut down a signal, as well as turn it on. Disrupting the activation—and deactivation—of G proteins can have dire consequences for a cell or organism. Consider cholera, for example. The disease is caused by a bacterium that multiplies in the human intestine, where it produces a protein called cholera toxin. This protein enters the cells that line the intestine and modifies the α subunit of a G protein called Gs—so named because it stimulates an enzyme called adenylyl cyclase, which we discuss shortly. The modification prevents Gs from hydrolyzing its bound GTP, thus locking the G protein in an active state, in which it continuously stimulates adenylyl cyclase. In intestinal cells, this stimulation causes a prolonged and excessive outflow of Cl– and water into the gut, resulting in catastrophic diarrhea and dehydration. The condition often leads to death unless urgent steps are taken to replace the lost water and ions.
A similar situation occurs in whooping cough (pertussis), a common respiratory infection against which infants are now routinely vaccinated. In this case, the disease-causing bacterium colonizes the lung, where it produces a protein called pertussis toxin. This protein alters the α subunit of a different type of G protein, called Gi because it inhibits adenylyl cyclase. In this case, however, modification by the toxin disables the G protein by locking it into its inactive GDP-bound state. Inhibiting Gi, like activating Gs, results in the prolonged and inappropriate activation of adenylyl cyclase, which, in this case, stimulates coughing. Both the diarrhea-producing effects of cholera toxin and the cough-provoking effects of pertussis toxin help the disease-causing bacteria spread from one host to another.
Some G Proteins Directly Regulate Ion Channels
There are about 20 different types of mammalian G proteins, each activated by a particular set of cell-surface receptors and dedicated to activating a particular set of target proteins. These target proteins are either enzymes or ion channels in the plasma membrane. Thus, the binding of an extracellular signal molecule to a GPCR leads to changes in the activities of a specific subset of the possible target proteins in the plasma membrane, producing a response that is appropriate for that signal and that type of cell.
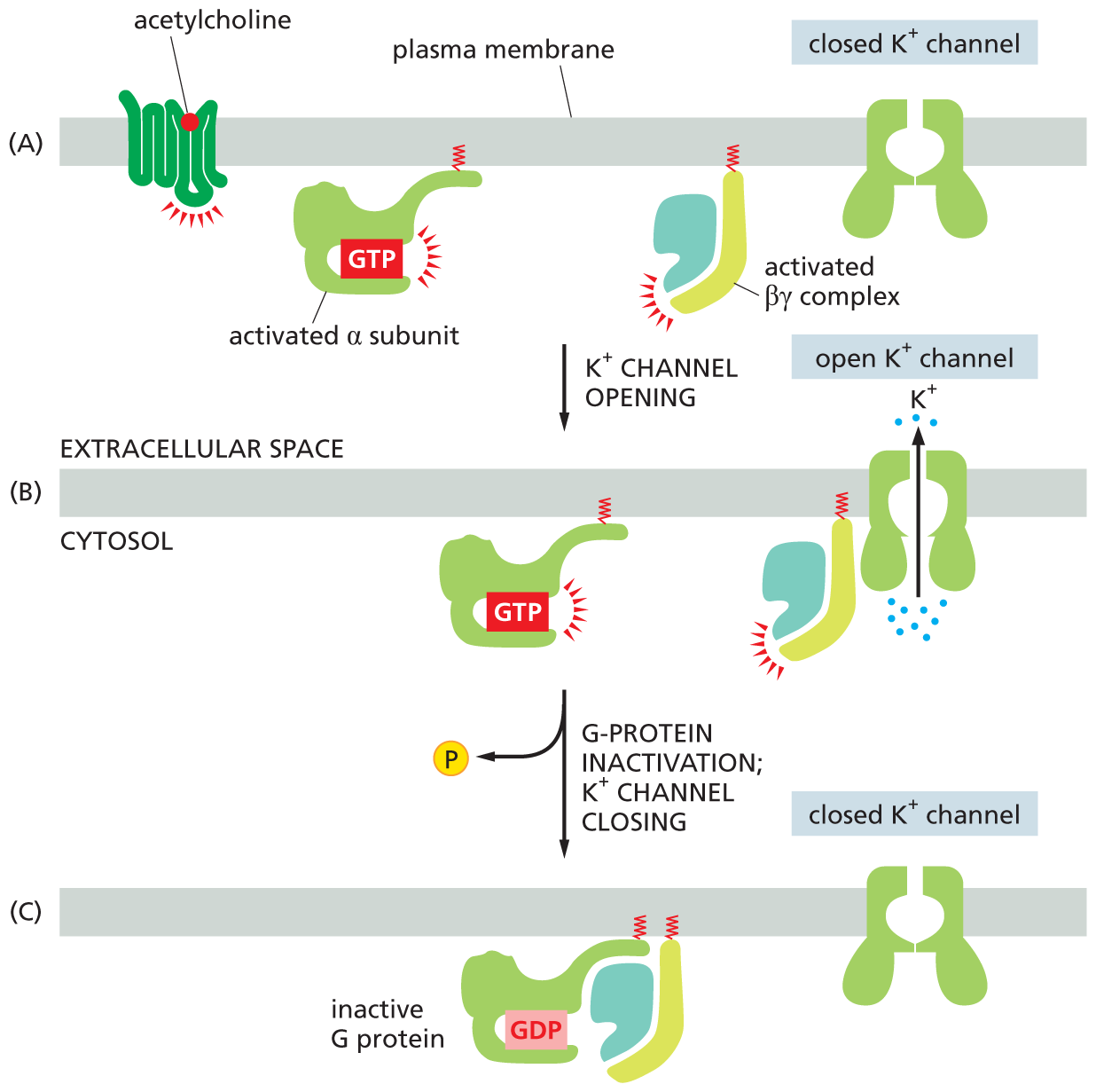
Some G proteins directly regulate the activity of ion channels. The heartbeat in animals is controlled by two sets of nerves: one speeds the heart up, the other slows it down. The nerves that signal a slowdown in heartbeat do so by releasing acetylcholine (see Figure 16–5A), which binds to a GPCR on the surface of the heart pacemaker cells. In this case, when the GPCR activates the G protein, the βγ complex binds to the intracellular face of a K+ channel in the plasma membrane of the pacemaker cell, forcing the ion channel into an open conformation (Figure 16–17A and B). This channel opening slows the heart rate by increasing the plasma membrane’s permeability to K+, which makes it more difficult to electrically activate, as explained in Chapter 12. The original signal is terminated—and the K+ channel recloses—when the α subunit inactivates itself by hydrolyzing its bound GTP, returning the G protein to its inactive state (Figure 16–17C).
More information
Three illustrations show the function of a G protein in heart pacemaker cells. Illustration A shows a receptor protein with an acetylcholine signal molecule attached to the plasma membrane. An activated alpha subunit with G T P and an activated beta gamma complex are shown bound to the plasma membrane on the cytosol side. A closed potassium ion channel is shown spanning the plasma membrane. The potassium ion channel opens, which leads to illustration B.
Illustration B shows potassium ions moving out from the cytosol to the extracellular space through the open potassium ion channel, with the activated beta gamma complex next to the potassium ion channel. Leading into illustration C is G protein inactivation, K plus channel closing, with a phosphate being removed from G T P via hydrolysis. Illustration C shows a closed potassium ion channel attached the plasma membrane. The inactive G protein comprising the alpha subunit with G D P, beta, and gamma subunits remains attached to the plasma membrane.
Figure 16–17A G protein directly couples receptor activation to the opening of K+ channels in the plasma membrane of heart pacemaker cells. (A) Binding of the neurotransmitter acetylcholine to its GPCR on heart cells results in the activation of the G protein, Gi. (B) The activated βγ complex directly opens a K+ channel in the plasma membrane, increasing its permeability to K+ and thereby making the membrane harder to activate and slowing the heart rate. (C) Inactivation of the α subunit by hydrolysis of its bound GTP returns the G protein to its inactive state, allowing the K+ channel to close.
Many G Proteins Activate Membrane-bound Enzymes That Produce Small Messenger Molecules
When G proteins interact with ion channels they cause an immediate change in the state and behavior of the cell. The interaction of activated G proteins with enzymes, in contrast, has consequences that are less rapid and more complex, as they lead to the production of additional intracellular signaling molecules. The two most frequent target enzymes for G proteins are adenylyl cyclase, which produces a small molecule called cyclic AMP, and phospholipase C, which generates small molecules called inositol trisphosphate and diacylglycerol. Inositol trisphosphate, in turn, promotes the accumulation of cytosolic Ca2+—yet another intracellular signaling molecule.
Adenylyl cyclase and phospholipase C are activated by different types of G proteins, allowing cells to couple the production of the small molecules to different extracellular signals. Although the coupling may be either stimulatory or inhibitory—as we saw in our discussion of the actions of cholera toxin and pertussis toxin—we concentrate here on G proteins that stimulate enzyme activity.
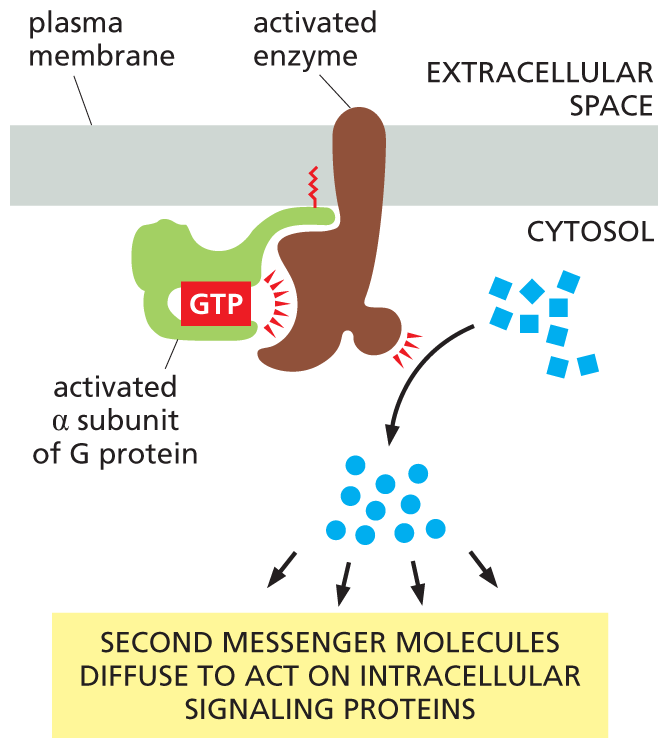
The small molecules generated by these enzymes are often called second messengers—the “first messengers” being the extracellular signals that activated the enzymes in the first place. Once activated, the enzymes generate large quantities of second messengers, which rapidly diffuse away from their source, thereby amplifying and spreading the intracellular signal (Figure 16–18).
More information
An illustration shows the effect on enzymes activated by G protein. An activated G protein alpha subunit bound to G T P is shown attached to the plasma membrane, and activates an enzyme bound to the plasma membrane. The activated enzyme produces messenger molecules in the cytosol. These messenger molecules lead to a second set of messenger molecules. Text reads, second messenger molecules diffuse to act on intracellular signaling proteins.
Figure 16–18Enzymes activated by G proteins increase the concentrations of small intracellular signaling molecules. Because each activated enzyme generates many molecules of these second messengers, the signal is greatly amplified at this step in the pathway (see Figure 16–28). The signal is relayed onward by the second messenger molecules, which bind to specific signaling proteins in the cell and influence their activity.
Different second messenger molecules produce different responses. We first examine the consequences of an increase in the cytosolic concentration of cyclic AMP. This will take us along one of the main types of signaling pathways that lead from the activation of GPCRs. We then discuss the actions of three other second messenger molecules—inositol trisphosphate, diacylglycerol, and Ca2+—which will lead us along a different signaling route.
The Cyclic AMP Signaling Pathway Can Activate Enzymes and Turn On Genes
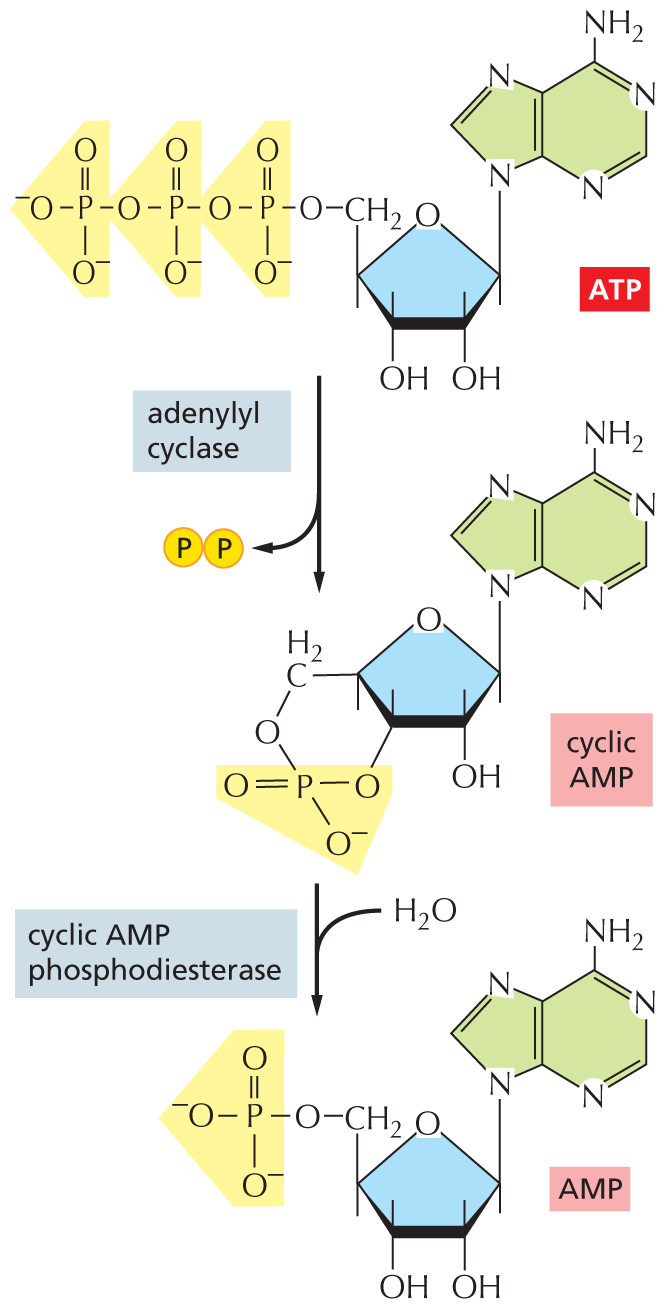
Many extracellular signals acting via GPCRs affect the activity of the enzyme adenylyl cyclase and thus alter the intracellular concentration of the second messenger molecule cyclic AMP. Most commonly, the activated G protein α subunit switches on the adenylyl cyclase, causing a dramatic and sudden increase in the synthesis of cyclic AMP from ATP (which is always present in the cell). To help terminate the signal, a second enzyme, called cyclic AMP phosphodiesterase, rapidly converts cyclic AMP to ordinary AMP (Figure 16–19). One way that caffeine acts as a stimulant is by inhibiting this phosphodiesterase in the nervous system, blocking cyclic AMP degradation and thereby keeping the concentration of this second messenger high.
More information
An illustration shows the formation and degradation of cyclic A M P. The chemical structures of the reactants are shown. The structure of A T P shows a ribose ring bonded to a linear chain of three phosphate groups on the left and an adenine ring at the top right. Two phosphate groups are removed from A T P via adenylyl cyclase. This forms the structure of cyclic A M P, which differs in having only one phosphate group instead of three, and the phosphate group is bound to the ribose ring at the center of A T P. In a condensation reaction (with the addition of water), cyclic A M P phosphodiesterase breaks the bond of the single phosphate group the the ribose ring of the cyclic A M P, creating A M P.
Figure 16–19Cyclic AMP is synthesized by adenylyl cyclase and degraded by cyclic AMP phosphodiesterase. Cyclic AMP (often abbreviated cAMP) is formed from ATP by a cyclization reaction that removes two phosphate groups from ATP and joins the remaining phosphate group to the sugar part of the AMP molecule. The degradation reaction breaks this new bond, forming AMP.
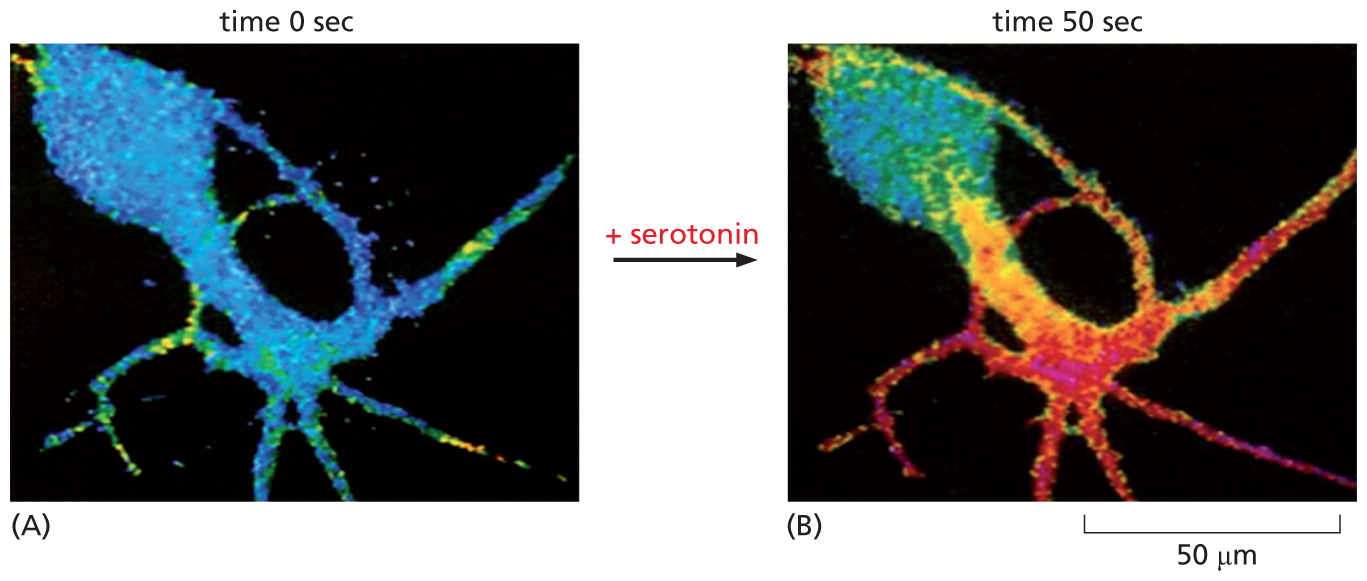
Cyclic AMP phosphodiesterase is continuously active inside the cell. Because it eliminates cyclic AMP so quickly, the cytosolic concentration of this second messenger can change rapidly in response to extracellular signals, rising or falling tenfold in a matter of seconds (Figure 16–20). Cyclic AMP is water-soluble, so it can, in some cases, carry the signal throughout the cell, traveling from the site on the membrane where it is synthesized to interact with proteins located in the cytosol, in the nucleus, or on other organelles.
More information
Two fluorescence micrographs show a nerve cell responding to serotonin. The first micrograph shows a nerve cell at 0 seconds, colored blue with small sections of green and yellow. The color blue reflects a low concentration of cyclic A M P, with yellow and green representing areas of the cell with higher concentrations of cyclic A M P. Serotonin is added between micrograph A and micrograph B. Micrograph B shows the cell 50 seconds following the addition of serotonin, on a scale of fifty micrometers. The nerve cell is red and purple at the dendrites, indicating a very high concentration of cyclic A M P.
Figure 16–20The concentration of cyclic AMP rises rapidly in response to an extracellular signal. A nerve cell in culture responds to the binding of the neurotransmitter serotonin to a GPCR by synthesizing cyclic AMP. The concentration of intracellular cyclic AMP is monitored by injecting into the cell a fluorescent protein whose fluorescence changes when it binds cyclic AMP. Blue indicates a low level of cyclic AMP, yellow an intermediate level, and red a high level. (A) In the resting cell, the cyclic AMP concentration is about 5 × 10–8 M. (B) Less than 1 minute after adding serotonin to the culture medium, the intracellular concentration of cyclic AMP has risen more than twentyfold (to >10–6 M) in the parts of the cell where the serotonin receptors are concentrated. (From B.J. Bacskai et al., Science 260:222–226, 1993.)
Cyclic AMP exerts most of its effects by activating the enzyme cyclic-AMP-dependent protein kinase (PKA). This enzyme is normally held inactive in a complex with a regulatory protein. The binding of cyclic AMP to the regulatory protein forces a conformational change that releases the inhibition and unleashes the active kinase. Activated PKA then catalyzes the phosphorylation of particular serines or threonines on specific intracellular proteins, thus altering the activity of these target proteins. In different cell types, different sets of proteins are available to be phosphorylated, which largely explains why the effects of cyclic AMP vary with the type of target cell.
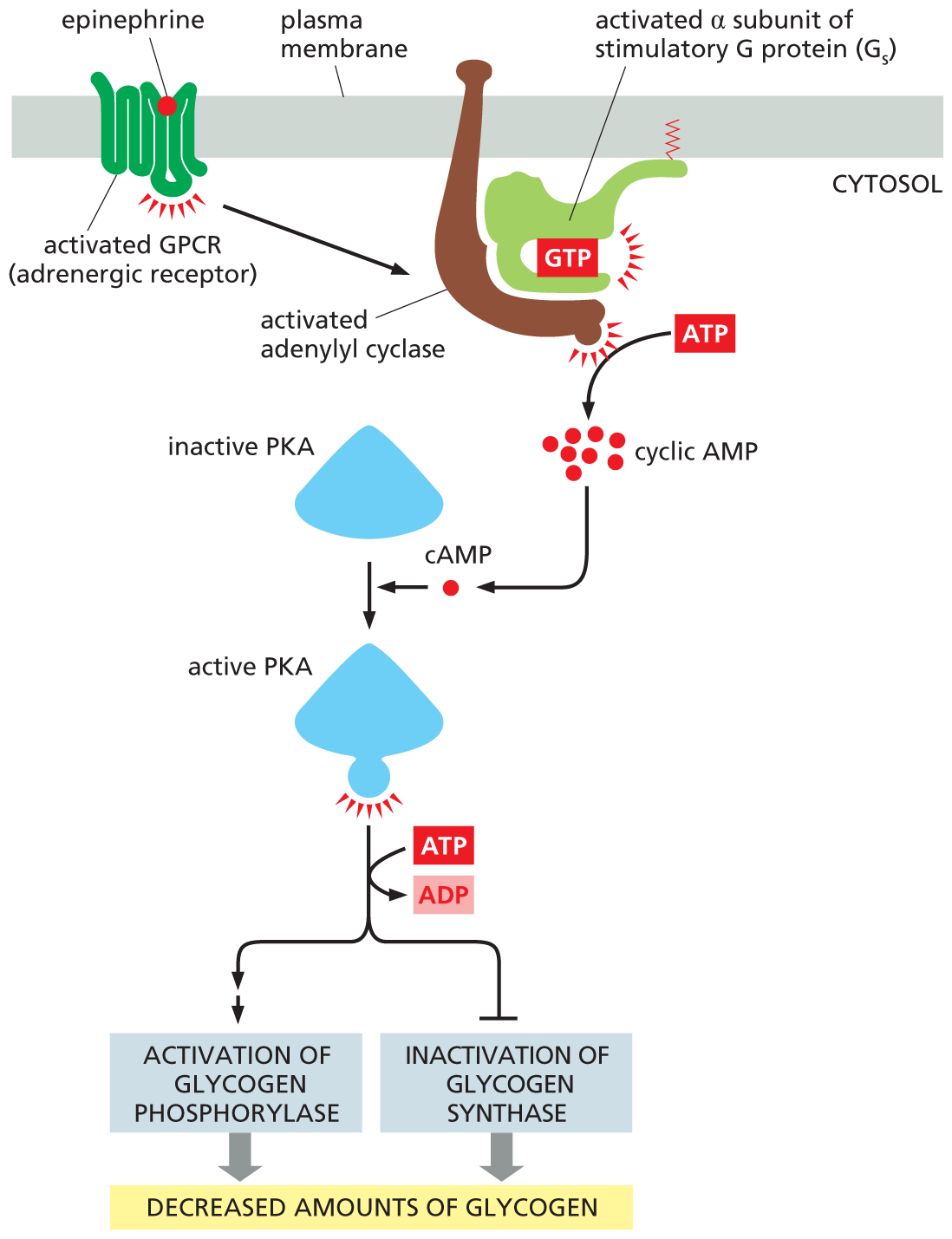
Many kinds of cell responses are mediated by cyclic AMP; a few are listed in Table 16–3. As the table shows, different target cells respond very differently to extracellular signals that change intracellular cyclic AMP concentrations. When we are frightened or excited, for example, the adrenal gland releases the hormone epinephrine (also called adrenaline), which circulates in the bloodstream and binds to a class of GPCRs called adrenergic receptors (see Figure 16–14B), which are present on many types of cells. The consequences of epinephrine binding vary from one cell type to another, but all the cell responses help prepare the body for sudden action. In skeletal muscle, for instance, epinephrine increases intracellular cyclic AMP, causing a decrease in the amount of glycogen—the polymerized storage form of glucose. It does so by activating PKA, which leads to both the activation of an enzyme that promotes glycogen breakdown and the inhibition of an enzyme that drives glycogen synthesis (Figure 16–21). By stimulating glycogen breakdown and inhibiting its synthesis, the increase in cyclic AMP maximizes the amount of glucose available as fuel for anticipated muscular activity. Epinephrine also acts on fat cells, stimulating the breakdown of fat to fatty acids. These fatty acids can then be exported to fuel ATP production in other cells.
TABLE 16–3 SOME CELL RESPONSES MEDIATED BY CYCLIC AMP
A flow diagram shows how epinephrine stimulates glycogen breakdown in skeletal muscle cells. A schematic shows an activated G P C R (adrenergic receptor) bound to the plasma membrane. A G T P bound activated alpha-subunit of stimulatory G protein (G subscript s), bound to an activated adenylyl cyclase, is also attached to the plasma membrane. A T P forms cyclic A M P. A c A M P molecule combines with inactive P K A to form an active P K A. Active P K A, using the energy from A T P being converted to A D P, leads to activation of glycogen phosphorylase and inactivation of glycogen synthase. Both pathways result in decreased amounts of glycogen.
Figure 16–21Epinephrine decreases glycogen levels in skeletal muscle cells. The hormone activates a GPCR, which turns on a G protein (Gs) that activates adenylyl cyclase to boost the production of cyclic AMP. The increase in cyclic AMP activates PKA, which phosphorylates and activates another kinase (not shown) that then phosphorylates and activates glycogen phosphorylase—the enzyme that breaks down glycogen (see Figure 13–22). In addition, PKA phosphorylates and inactivates glycogen synthase, the enzyme that makes glycogen. Because these reactions do not involve changes in gene transcription or new protein synthesis, they occur rapidly.
QUESTION 16–4
In cells, cyclic AMP is broken down rapidly. Why does this allow for rapid signaling?
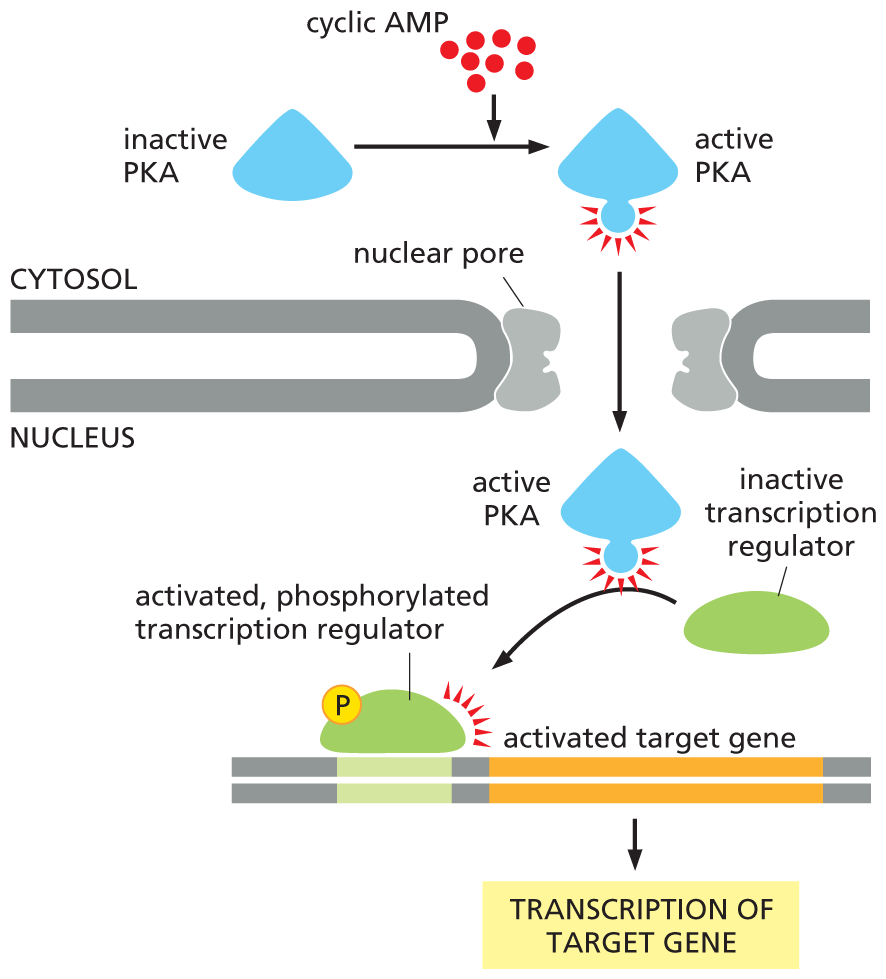
In some cases, the effects of increasing cyclic AMP are rapid; in skeletal muscle, for example, glycogen breakdown occurs within seconds of epinephrine binding to its receptor (see Figure 16–21). In other cases, cyclic AMP responses involve changes in gene expression that take minutes or hours to develop. In these slow responses, PKA typically phosphorylates transcription regulators, proteins that activate the transcription of selected genes (as discussed in Chapter 8). For example, an increase in cyclic AMP in certain neurons in the brain controls the production of proteins involved in some forms of learning. Figure 16–22 illustrates how a rise in intracellular cyclic AMP can activate the transcription of a target gene in the nucleus.
More information
A flow diagram shows how a rise in intracellular cyclic A M P can activate gene transcription. In the first step, cyclic A M P aids the conversion of inactive P K A to active P K A in the cytosol. Active P K A enters the nucleus through a nuclear pore and converts an inactive transcription regulator to an activated, phosphorylated transcription regulator, which activates the target gene. This leads to the transcription of the target gene in the final step.
Figure 16–22A rise in intracellular cyclic AMP can activate gene transcription. PKA, activated by a rise in intracellular cyclic AMP, can enter the nucleus and phosphorylate specific transcription regulators. Once phosphorylated, these proteins stimulate the transcription of a whole set of target genes (Movie 16.3). This type of signaling pathway controls many processes in cells, ranging from hormone synthesis in endocrine cells to the production of proteins involved in long-term memory in the brain. Activated PKA can also phosphorylate and thereby regulate other proteins and enzymes in the cytosol, as shown in Figure 16–21.
The Inositol Phospholipid Pathway Triggers a Rise in Intracellular Ca2+
We now turn to the other enzyme-mediated signaling pathway that leads from GPCRs: the pathway that begins with the activation of the membrane-bound enzyme phospholipase C. GPCRs that activate phospholipase C (rather than adenylyl cyclase) exert their effects through a G protein called Gq. Examples of signal molecules that act through phospholipase C are given in Table 16–4.
TABLE 16–4 SOME CELL RESPONSES MEDIATED BY PHOSPHOLIPASE C ACTIVATION
Signal Molecule
Target Tissue
Major Response
Vasopressin (a peptide hormone)
Liver
Glycogen breakdown
Acetylcholine
Pancreas
Secretion of amylase (a digestive enzyme)
Acetylcholine
Skeletal muscle
Contraction
Thrombin (a proteolytic enzyme)
Blood platelets
Aggregation
Once activated, phospholipase C propagates the signal by cleaving a lipid molecule that is a component of the plasma membrane. The molecule is an inositol phospholipid (a phospholipid with the sugar inositol attached to its head) that is present in small quantities in the cytosolic leaflet of the membrane lipid bilayer (see Figure 11–19). Because of the involvement of this phospholipid, the signaling pathway that begins with the activation of phospholipase C is often referred to as the inositol phospholipid pathway. It operates in almost all eukaryotic cells and regulates a large number of different effector proteins.
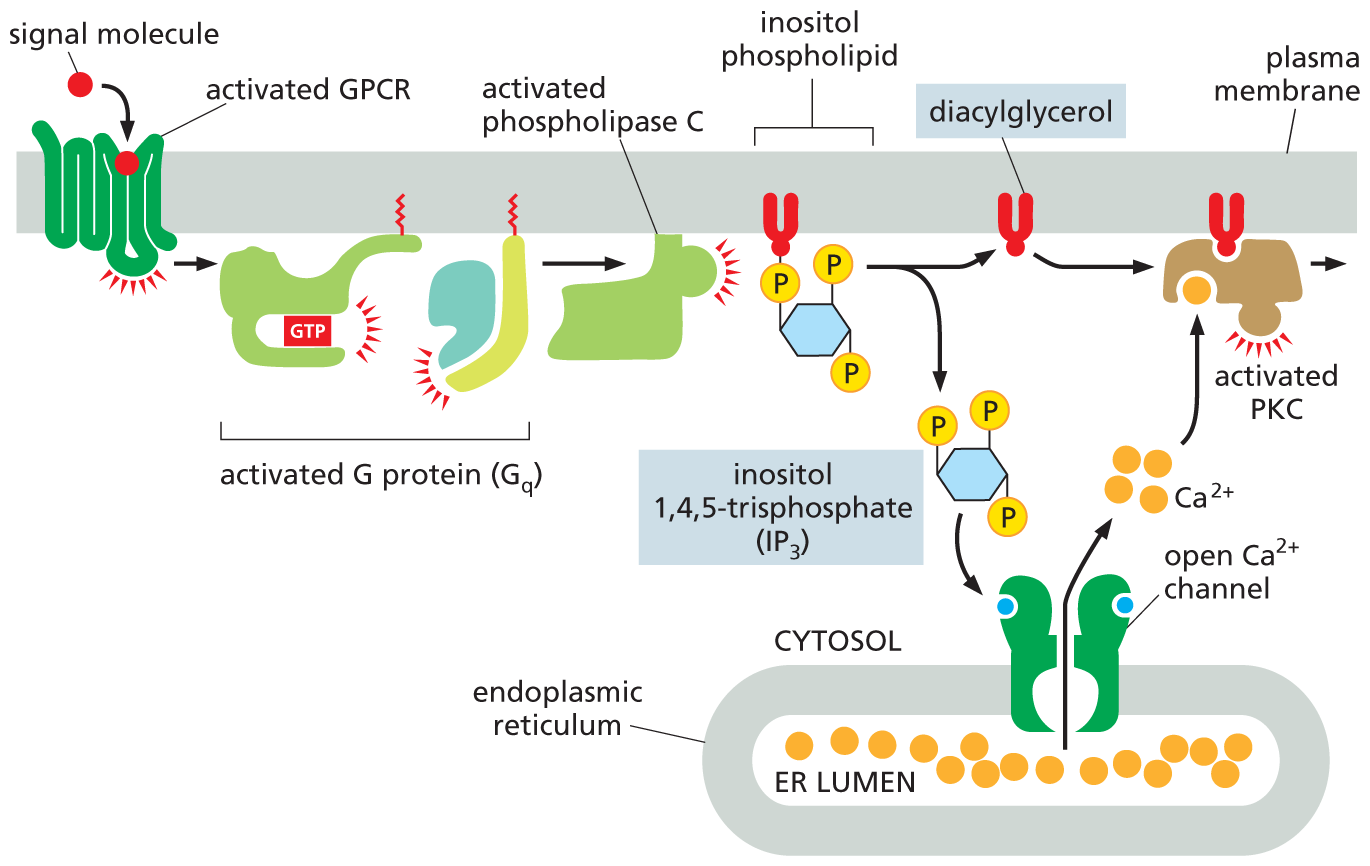
The cleavage of a membrane inositol phospholipid by phospholipase C generates two second messenger molecules: inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG). Both molecules play a crucial part in relaying the signal (Figure 16–23).
More information
An illustration shows how two signaling pathways are activated by phospholipase C. A signal molecule activates a G P C R which in turn activates a G protein (G subscript q) comprising a G T P bound alpha subunit and a beta-gamma complex. This activated G protein activates phospholipase C which hydrolyzes a membrane bound inositol phospholipid, forming diacylglycerol and inositol 1,4,5-triphosphate (I P subscript three). Inositol 1,4,5-triphosphate (I P subscript 3) triggers the release of ionized calcium (C A 2 plus) from an open ionized calcium channel on the the endoplasmic reticulum. C A 2 plus ions from the E R lumen exit the E R through the open C a 2 plus channel into the cytosol. The C a 2 plus ions and diacylglycerol bind to P K C, leading to an activated P K C bound to the plasma membrane.
Figure 16–23Phospholipase C activates two signaling pathways. Two messenger molecules are produced when a membrane inositol phospholipid is hydrolyzed by activated phospholipase C. Inositol 1,4,5-trisphosphate (IP3) diffuses through the cytosol and triggers the release of Ca2+ from the ER by binding to and opening special Ca2+ channels in the ER membrane. The large electrochemical gradient for Ca2+ across this membrane causes Ca2+ to rush out of the ER and into the cytosol. Diacylglycerol remains in the plasma membrane and, together with Ca2+, helps activate the enzyme protein kinase C (PKC), which is recruited from the cytosol to the cytosolic face of the plasma membrane. PKC then phosphorylates its own set of intracellular proteins, further propagating the signal (Movie 16.4). At the start of the pathway, both the α subunit and the βγ complex of the G protein Gq are involved in activating phospholipase C.
IP3 is a water-soluble sugar phosphate that is released into the cytosol; there it binds to and opens Ca2+ channels that are embedded in the endoplasmic reticulum (ER) membrane. Ca2+ stored inside the ER rushes out into the cytosol through these open channels, causing a sharp rise in the cytosolic concentration of free Ca2+, which is normally kept very low. This Ca2+ in turn signals to other proteins, as we discuss shortly.
Diacylglycerol is a lipid that remains embedded in the plasma membrane after it is produced by phospholipase C; there, it helps recruit and activate a protein kinase, which translocates from the cytosol to the plasma membrane. This enzyme is called protein kinase C (PKC) because it also needs to bind Ca2+ to become active (see Figure 16–23). Once activated, PKC phosphorylates a set of intracellular proteins that varies depending on the cell type.
A Ca2+ Signal Triggers Many Biological Processes
QUESTION 16–5
Why do you suppose cells have evolved intracellular Ca2+ stores for signaling even though there is abundant extracellular Ca2+?
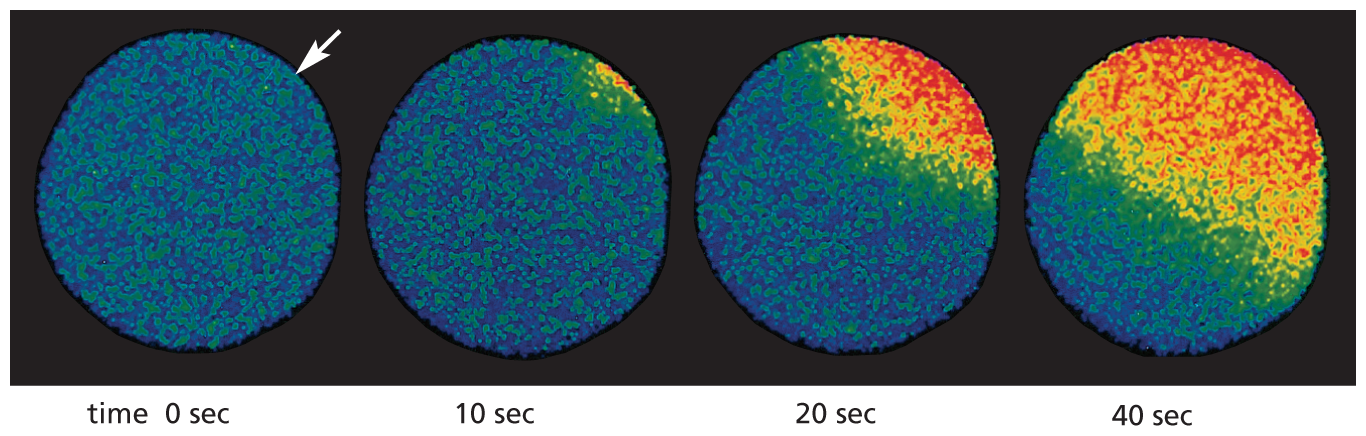
Ca2+ has such an important and widespread role as an intracellular messenger that we will digress to consider its functions more generally. A surge in the cytosolic concentration of free Ca2+ is triggered by many kinds of cell stimuli, not only those that act through GPCRs. When a sperm fertilizes an egg cell, for example, Ca2+ channels open, and the resulting rise in cytosolic Ca2+ triggers the egg to start development (Figure 16–24); for muscle cells, a signal from a nerve triggers a rise in cytosolic Ca2+ that initiates muscle contraction (discussed in Chapter 17; see Figure 17–45); and in many secretory cells, including nerve cells, Ca2+ triggers secretion (discussed in Chapter 12; see Figure 12–42). Ca2+ stimulates all these responses by binding to and influencing the activity of various Ca2+-responsive proteins.
More information
A set of four fluorescence micrographs show ionized calcium concentrations over time following egg fertilization. At 0 seconds, the sperm enters into the egg. The egg is a grainy blue and green, indicating low ionized calcium (C A 2 plus). An arrow is at the top right corner of the micrograph, showing the site of sperm entry. After 10 seconds, a small orange area, indicating higher ionized calcium concentration, appears closest to the site of sperm entry. After 20 seconds, the area spreads to one-fourth of the egg surface. At 40 seconds, half of the egg is red and orange.
Figure 16–24Fertilization of an egg by a sperm triggers an increase in cytosolic Ca2+ in the egg. This starfish egg was injected with a Ca2+-sensitive fluorescent dye before it was fertilized. When a sperm enters the egg, a wave of cytosolic Ca2+ (red)—released from the ER—sweeps across the egg from the site of sperm entry (arrow). This Ca2+ wave provokes a change in the egg surface, preventing entry of other sperm, and it also initiates embryonic development. To catch this Ca2+ wave, go to Movie 16.5. (Adapted from S.A. Stricker et al., Dev. Biol. 166:34–58, 1994.)
The concentration of free Ca2+ in the cytosol of an unstimulated cell is extremely low (10–7 M) compared with its concentration in the extracellular fluid (about 10–3 M) and in the ER. These differences are maintained by membrane-embedded Ca2+ pumps that actively remove Ca2+ from the cytosol, sending it either into the ER or across the plasma membrane and out of the cell. As a result, a steep electrochemical gradient of Ca2+ exists across both the ER membrane and the plasma membrane (discussed in Chapter 12). When a signal transiently opens Ca2+ channels in either of these membranes, Ca2+ rushes down its electrochemical gradient into the cytosol, where it triggers changes in Ca2+-responsive proteins. The same Ca2+ pumps that normally operate to keep cytosolic Ca2+ concentrations low also help to terminate the Ca2+ signal.
The effects of Ca2+ in the cytosol are largely indirect, in that they are mediated through the interaction of Ca2+ with various kinds of Ca2+-responsive proteins. The most widespread and common of these is calmodulin, which is present in the cytosol of all eukaryotic cells that have been examined, including those of plants, fungi, and protozoa. When Ca2+ binds to calmodulin, the protein undergoes a conformational change that enables it to interact with a wide range of target proteins in the cell, altering their activities (Figure 16–25). One particularly important class of targets for calmodulin is the Ca2+/calmodulin-dependent protein kinases (CaM-kinases). When these kinases are activated by binding to calmodulin complexed with Ca2+, they influence other processes in the cell by phosphorylating selected proteins. In the mammalian brain, for example, a neuron-specific CaM-kinase is abundant at synapses, where it is thought to play an important part in some forms of learning and memory. This CaM-kinase is activated by the pulses of Ca2+ signals that occur during neural activity, and mutant mice that lack the kinase show a marked inability to remember where things are.
More information
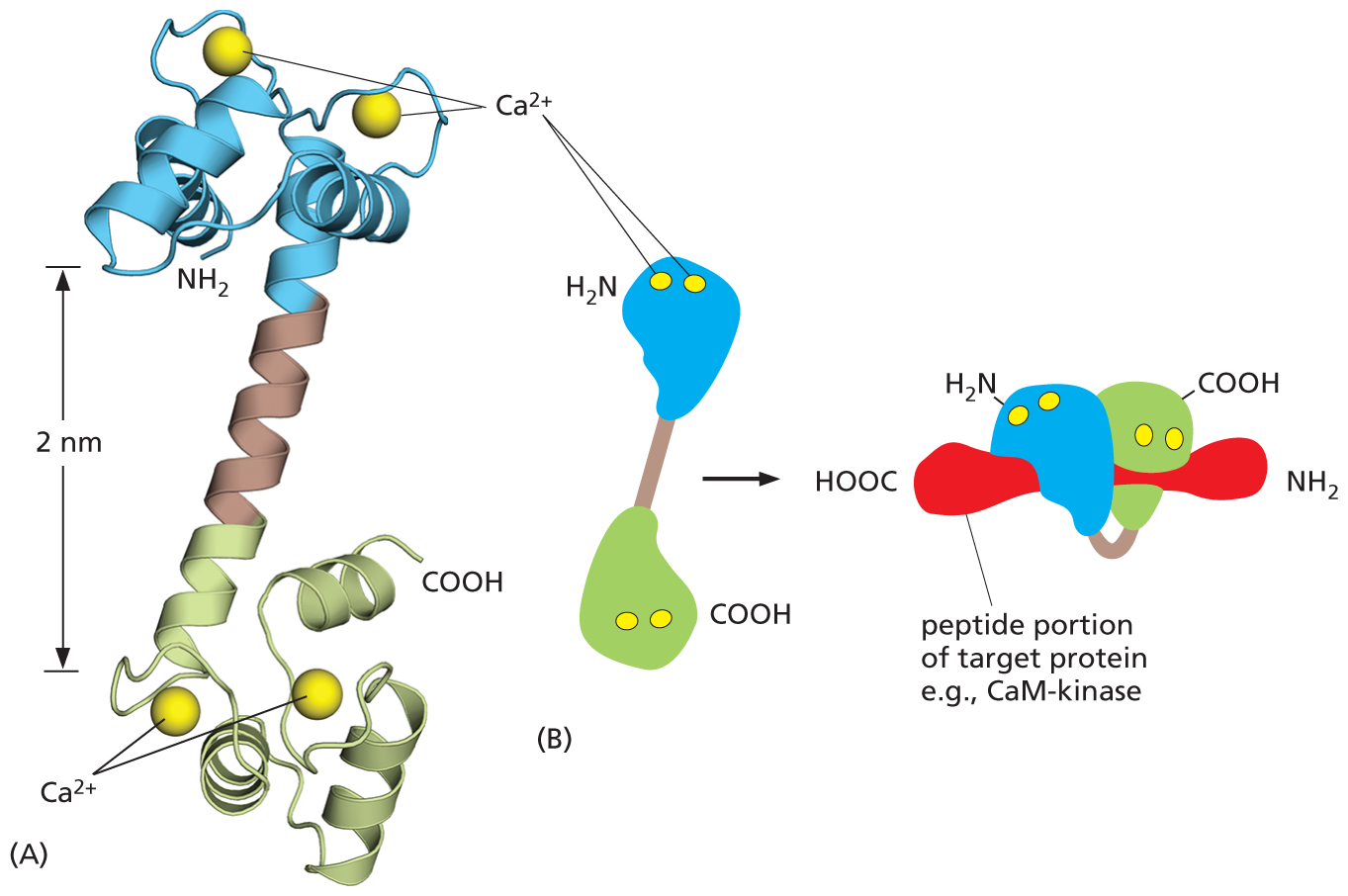
Two illustrations show the shape of calmodulin protein after binding to calcium ions. Illustration A shows a ribbon model of calmodulin. Two C a 2 plus ions are bound to alpha helices at the top and bottom that resemble a dumbbell. The helix on top has a N H subscript 2 terminus and the helix at the bottom has a C O O H terminus. The vertical distance between the top and the bottom part of the dumbbell reads 2 nanometers.
Illustration B shows a simplified model of calmodulin binding to a target protein. Calmodulin is dumbbell shaped. The top globular end has two C a 2 plus ions and a N H subscript 2 terminus. The bottom globular end has two C a 2 plus ions and a C O O H terminus. This structure leads to a target protein (example: C a M kinase) which is bound to calmodulin.
A horizontal peptide portion of a target protein is shown (such as C a M kinase), with calmodulin wrapped around it. The C O O H terminus of calmodulin is on the right side of the protein, closest to the N H subscript 2 terminus of the peptide portion of the target protein. The N H subscript 2 terminus of calmodulin is on the left, closest to the C O O H end of the peptide portion of the target protein. The alpha helix connecting the termini (arm of the dumbbell shape) is wrapped around the peptide portion of the target protein.
Figure 16–25Calcium binding changes the shape of the calmodulin protein. (A) Calmodulin has a dumbbell shape, with two globular ends connected by a long α helix. Each of the globular ends has two Ca2+-binding sites. (B) Simplified representation of the structure, showing the conformational changes that occur when Ca2+-bound calmodulin interacts with an isolated segment of a target protein (red). In this conformation, the α helix jackknifes to allow calmodulin to wrap around its target (Movie 16.6). (Dynamic Figure) The binding of calcium (yellow) triggers a conformational change that allows calmodulin to wrap around its target protein (red).
Some GPCR Signaling Pathways Generate a Dissolved Gas That Carries a Signal to Adjacent Cells
Second messengers like cyclic AMP and Ca2+ are hydrophilic, and they generally interact with target proteins within the cell in which they are generated. But some molecules produced in response to GPCR activation are small enough or hydrophobic enough to pass across the membrane and carry a signal directly to nearby cells. An important example is the gas nitric oxide (NO), which acts as a signaling molecule in many tissues. NO diffuses readily from its site of synthesis and slips into neighboring cells. The distance the gas diffuses is limited by its reaction with oxygen and water in the extracellular environment, which converts NO into nitrates and nitrites within seconds.
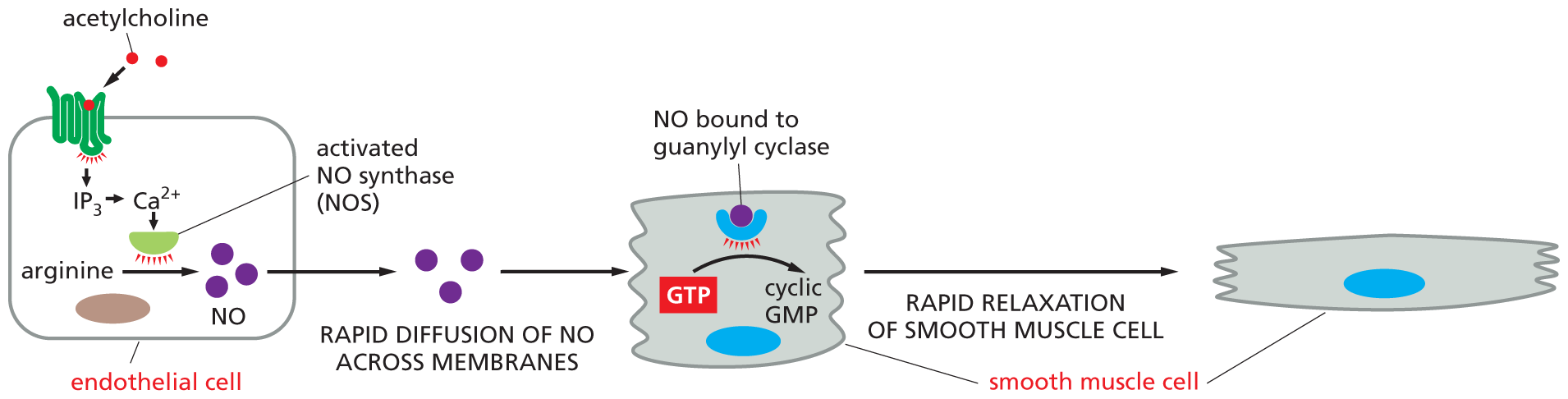
Endothelial cells—the flattened cells that line every blood vessel—release NO in response to acetylcholine secreted by nearby nerve endings. Acetylcholine binds to a GPCR on the endothelial cell surface, resulting in activation of Gq and the release of Ca2+ inside the cell (see Figure 16–23). Ca2+ then stimulates nitric oxide synthase, which produces NO from the amino acid arginine. This NO diffuses into smooth muscle cells in the adjacent vessel wall, causing the cells to relax; this relaxation allows the vessel to dilate, so that blood flows through it more freely (Figure 16–26). The effect of NO on blood vessels accounts for the action of nitroglycerin, which has been used for almost 100 years to treat patients with angina—pain caused by inadequate blood flow to the heart muscle. In the body, nitroglycerin is converted to NO, which rapidly relaxes blood vessels, thereby reducing the workload on the heart and decreasing the muscle’s need for oxygen-rich blood. Many nerve cells also use NO to signal neighboring cells: NO released by nerve terminals in the penis, for instance, acts as a local mediator to trigger the blood-vessel dilation responsible for penile erection.
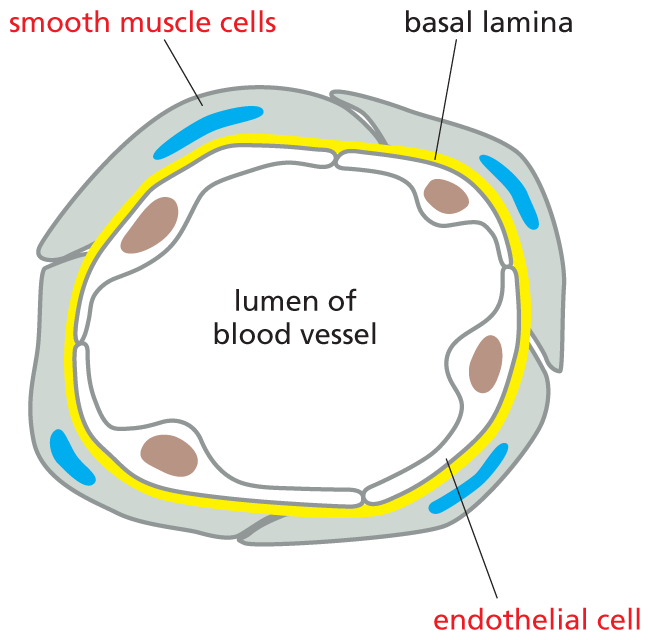
(A)
More information
Illustration A shows a cross section of a blood vessel. The cross section shows the basal lamina surrounded by a layer of smooth muscle cells. Inside the basal lamina, the lumen of the blood vessel is labeled.
(B)
More information
Illustration B shows the method by which smooth muscle cell relaxation occurs. An endothelial cell is shown. Acetylcholine binds to the G P C R on the surface of the endothelial cell and activates it. This activated protein leads to IP subscript 3, and then stimulates release of C a 2 plus ions. The ionized calcium activates N O S (N O synthase). N O S aids the conversion of arginine to N O molecules inside the endothelial cell. In the next step, rapid diffusion of N O across membranes occurs, and G T P is converted to cyclic G M P, by the action of N O bound guanylyl cyclase in the smooth muscle cell. This leads to rapid relaxation of the smooth muscle cell in the final step, and the final cell shown is wider than the smooth muscle cell in the second step.
Figure 16–26Nitric oxide (NO) triggers smooth muscle relaxation in a blood-vessel wall. (A) Simplified drawing showing a cross section of a blood vessel with endothelial cells lining its lumen and smooth muscle cells surrounding the outside of the vessel. (B) The neurotransmitter acetylcholine causes the blood vessel to dilate by binding to a GPCR on the surface of the endothelial cells, thereby activating a G protein, Gq, to trigger Ca2+ release (as illustrated in Figure 16–23). Ca2+ activates nitric oxide synthase, stimulating the production of NO. NO then diffuses out of the endothelial cells and into adjacent smooth muscle cells, where it regulates the activity of specific proteins, causing the muscle cells to relax. One key target protein that can be activated by NO in smooth muscle cells is guanylyl cyclase, which catalyzes the production of cyclic GMP from GTP. Note that NO gas is highly toxic when inhaled and should not be confused with nitrous oxide (N2O), also known as laughing gas, which is sometimes used as a sedative.
Inside many target cells, NO binds to and activates the enzyme guanylyl cyclase, stimulating the formation of cyclic GMP from the nucleotide GTP (see Figure 16–26B). Cyclic GMP, a second messenger similar in structure to cyclic AMP, is a key link in the NO signaling chain. The drug Viagra enhances penile erection by blocking the enzyme that degrades cyclic GMP, prolonging the NO signal.
GPCR-triggered Intracellular Signaling Pathways Can Achieve Astonishing Speed, Sensitivity, and Adaptability
It seems likely that the GPCR-based mechanism of signal reception, which arose early in evolution, has its origins in the basic and universal need of cells to sense and respond to their environment. Our ability to taste, smell, and see all depend on GPCRs. And though steps in the signaling pathways associated with GPCRs can take a long time (and many textbook pages) to describe, they often take only seconds to execute. Consider how quickly a thrill can make your heart race (when epinephrine stimulates the GPCRs in your cardiac pacemaker cells), or how fast the smell of food can make your mouth water (through the GPCRs for odors in your nose and the GPCRs for acetylcholine in salivary cells, which stimulate secretion). Among the fastest of all responses mediated by a GPCR, however, is the response of the eye to light: it takes only 20 msec for the most quickly responding photoreceptor cells of the retina (the cone photoreceptors, which are responsible for color vision in bright light) to produce their electrical response to a sudden flash of light.
This exceptional speed is achieved in spite of the necessity to relay the signal over the multiple steps of an intracellular signaling cascade. The quantitative details of GCPR-based visual signaling pathways have been most thoroughly analyzed for the rod photoreceptor cells in the eye, which are responsible for noncolor vision in dim light (Figure 16–27). In this photoreceptor cell, light is sensed by rhodopsin, a G-protein-coupled light receptor. Rhodopsin, when stimulated by light, activates a G protein called transducin. The activated α subunit of transducin then activates an intracellular signaling cascade that causes cation channels to close in the plasma membrane of the photoreceptor cell. This produces a change in the voltage across the cell membrane, which alters neurotransmitter release and ultimately leads to a nerve impulse being sent to the brain.
More information
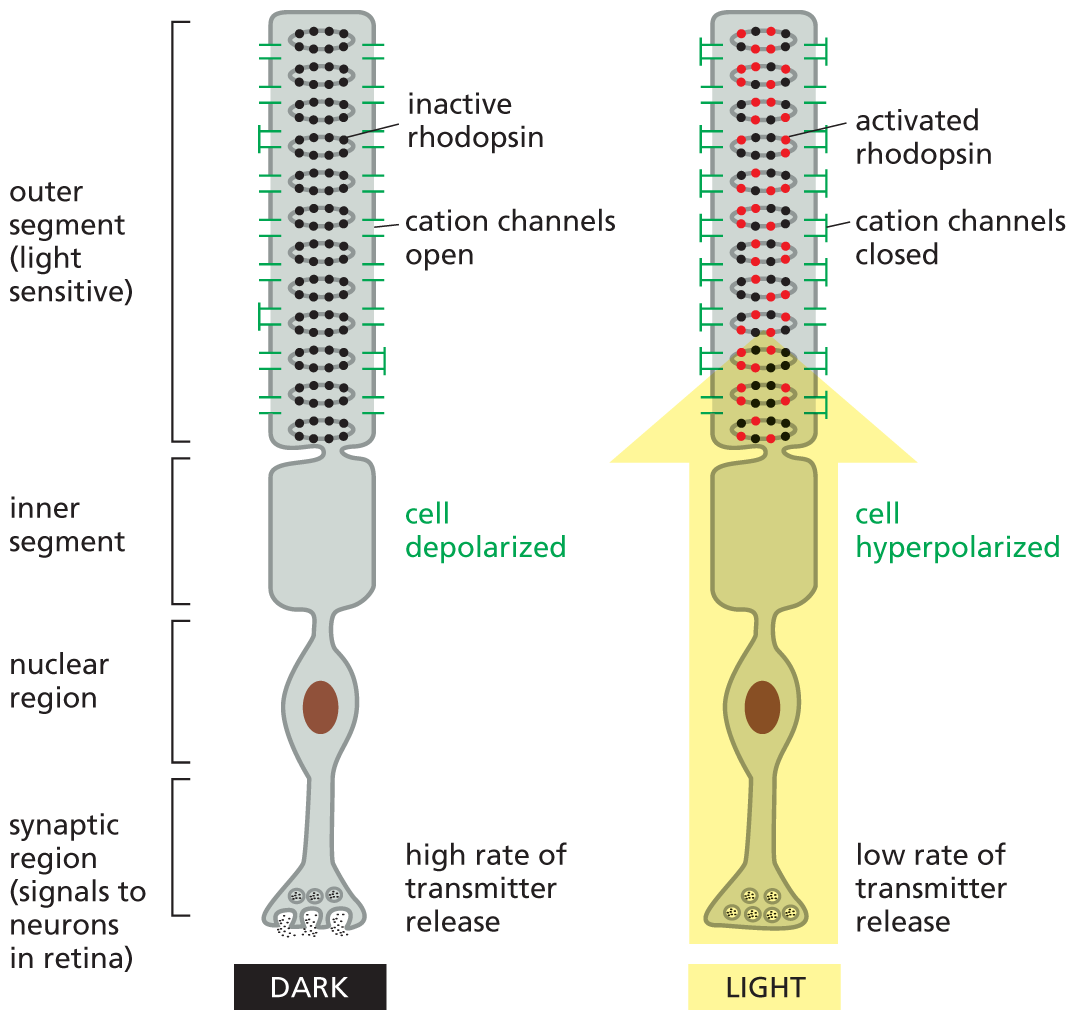
A schematic depicts a rod photoreceptor cell from the retina. The photoreceptor cell is shown in both light and dark conditions. The photoreceptor cell has a light sensitive outer segment, inner segment, nuclear region, and synaptic region that signals to neurons in the retina. In dark conditions, there is a high rate of transmitter release in the synaptic region, and the inner segment is depolarized. The outer segment has inactive rhodopsin, and cation channels are open. In light conditions, there is a low rate of transmitter release in the synaptic region, the inner segment is hyperpolarized, and the outer segment contains activated rhodopsin and cation channels are closed. An arrow extending from the synaptic region to the outer segment is shown in the cell during light conditions, symbolizing light.
Figure 16–27A rod photoreceptor cell from the retina is exquisitely sensitive to light. The light-absorbing rhodopsin proteins are embedded in many pancake-shaped vesicles (discs) of membrane inside the outer segment of the photoreceptor cell. When the rod cell is stimulated by light, a signal is relayed from the rhodopsin molecules in the discs, through the cytosol, to cation channels in the plasma membrane of the outer segment. These cation channels close in response to the cytosolic signal, producing a change in the membrane potential of the rod cell. By mechanisms similar to those that control neurotransmitter release in ordinary nerve cells, the change in membrane potential alters the rate of neurotransmitter release from the synaptic region of the cell. Released neurotransmitters act on retinal nerve cells that pass the signal on to the brain.
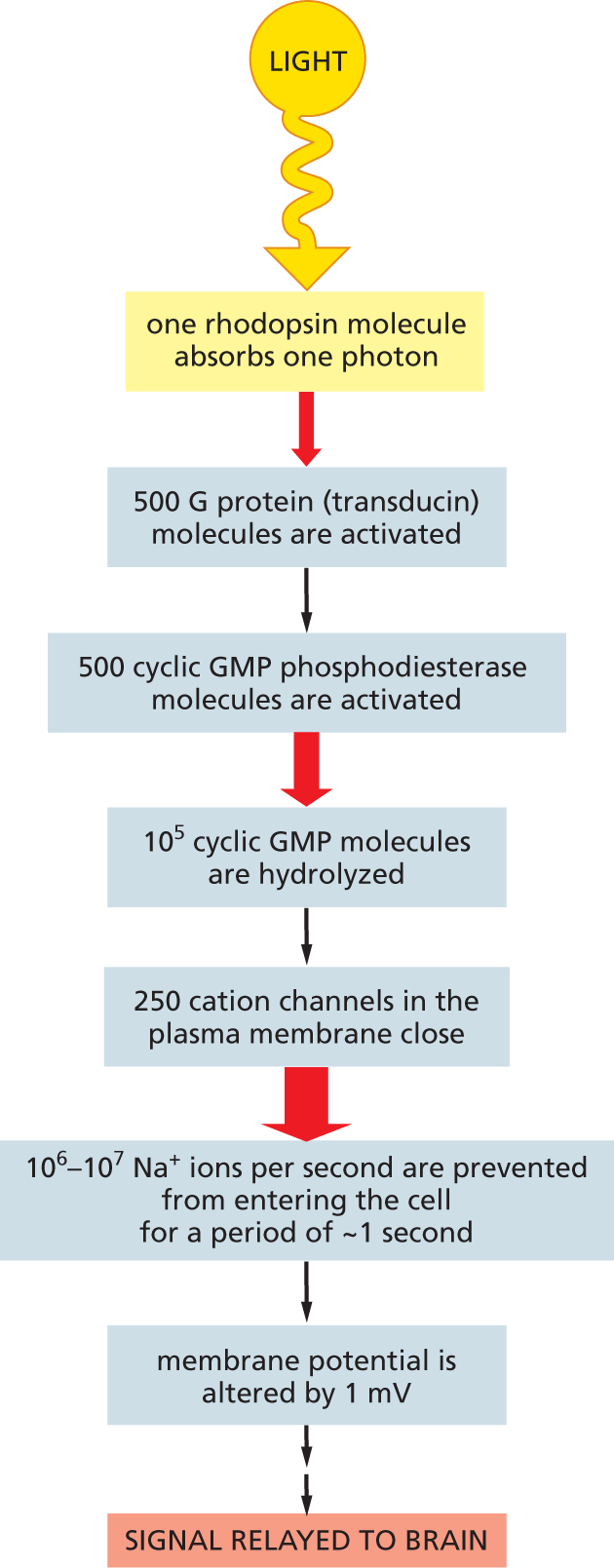
A key advantage of such multistep intracellular signaling cascades is that they allow spectacular amplification of the incoming signal (Figure 16–28). At the same time, they allow cells to adapt so as to be able to detect signals of widely varying intensity. When lighting conditions are dim, as on a moonless night, the signaling cascade boosts the signal tremendously: as few as a dozen photons absorbed across the entire retina will cause a perceptible signal to be delivered to the brain. In bright sunlight, when photons flood through each photoreceptor cell at a rate of billions per second, the signaling cascade will step down the amplification more than 10,000-fold, so that the photoreceptor cells are not overwhelmed and can still register increases and decreases in the strong light. This adjustment of sensitivity—a form of adaptation—depends on negative feedback: an intense response in the photoreceptor cell decreases the cytosolic Ca2+ concentration, inhibiting the enzymes responsible for signal amplification.
More information
A flow diagram depicts path of the light signal from photoreceptor cells to the brain. The steps are listed as follows:
Step 1: one rhodopsin molecule absorbs one photon.
Step 2: 500 G protein (transducin) molecules are activated.
Step 3: 500 cyclic G M P phosphodiesterase molecules are activated.
Step 4: 10 to the fifth power cyclic GMP molecules are hydrolyzed.
Step 6: 250 cation channels in the plasma membrane close.
Step 7: 10 to the sixth power to 10 to the seventh power N a plus ions per second are prevented from entering the cell for a period of approximately 1 second.
Step 8: membrane potential is altered by 1 millivolt.
Step 9: signal relayed to brain.
Figure 16–28The light-induced signaling cascade in rod photoreceptor cells greatly amplifies the light signal. When rod photoreceptors are adapted for dim light, the signal amplification is enormous. This light-sensing cascade functions as follows. In the absence of a light signal, the second messenger molecule cyclic GMP is continuously produced by guanylyl cyclase in the cytosol of the photoreceptor cell. The cyclic GMP then binds to cation channels in the photoreceptor cell plasma membrane, keeping them open. Activation of rhodopsin by light triggers the activation of the α subunit of a G protein called transducin. Transducin turns on an enzyme called cyclic GMP phosphodiesterase, which breaks down cyclic GMP to GMP (much as cyclic AMP phosphodiesterase breaks down cyclic AMP; see Figure 16–19). The sharp fall in the cytosolic concentration of cyclic GMP reduces the amount of cyclic GMP bound to the cation channels, which therefore close. Closing these channels decreases the influx of Na+, thereby altering the voltage gradient (membrane potential) across the plasma membrane and, ultimately, the rate of neurotransmitter release, as described in Chapter 12. The red arrows indicate the steps at which amplification occurs, with the thickness of the arrow roughly indicating the magnitude of the amplification.
Adaptation can also take place at the level of the receptor itself. GPCRs can be inactivated such that they no longer interact with G proteins; they can also be removed from the cell surface and temporarily sequestered so they can no longer interact with signal molecules—or routed to lysosomes for destruction (as described in Chapter 15).
Adaptation frequently occurs in intracellular signaling pathways that respond to extracellular signal molecules, allowing cells to respond to fluctuations in the concentration of such molecules regardless of whether they are present in small or large amounts. By taking advantage of mechanisms that promote amplification and adaptation, intracellular signaling pathways allow cells to respond equally well to the signaling equivalents of shouts and whispers. Of course, GPCRs are not the only receptors that activate intracellular signaling cascades. We now turn to another major class of cell-surface receptors—enzyme-coupled receptors—which play a key part in controlling cell numbers, cell differentiation, and cell movement in multicellular animals, especially during development.
A membrane-bound GTP-binding protein involved in intracellular signaling; composed of three subunits, this intermediary is usually activated by the binding of a hormone or other ligand to a transmembrane receptor.
Minor lipid component of plasma membranes that plays a part in signal transduction in eukaryotic cells; cleavage yields two small messenger molecules, IP3 and diacylglycerol.
Small messenger molecule produced by the cleavage of membrane inositol phospholipids in response to extracellular signals. Helps activate protein kinase C.
Adjustment of sensitivity following repeated stimulation; allows a cell or organism to register small changes in a signal despite a high background level of stimulation.
Cell-surface receptor that associates with an intracellular trimeric GTP-binding protein (G protein) after activation by an extracellular ligand. These receptors are embedded in the membrane by seven transmembrane α helices.
Enzyme that phosphorylates target proteins in response to an increase in Ca2+ ion concentration through its interaction with the Ca2+-binding protein calmodulin.
Small intracellular signaling molecule that triggers the release of Ca2+ from the endoplasmic reticulum into the cytosol; produced when a signal molecule activates a membrane-bound protein called phospholipase C.
Endnotes
Although all of the signal molecules listed here are hormones, some responses to local mediators and to neurotransmitters are also mediated by cyclic AMP.Return to reference *