Matter is made of combinations of elements—substances such as hydrogen or carbon that cannot be broken down or interconverted by chemical means. The smallest particle of an element that still retains its distinctive chemical properties is an atom. The characteristics of substances other than pure elements—including the materials from which living cells are made—depend on which atoms they contain and the way that these atoms are linked together in groups to form molecules. To understand living organisms, therefore, it is crucial to understand the chemical bonds that hold atoms together in molecules.
Cells Are Made of Relatively Few Types of Atoms
More information
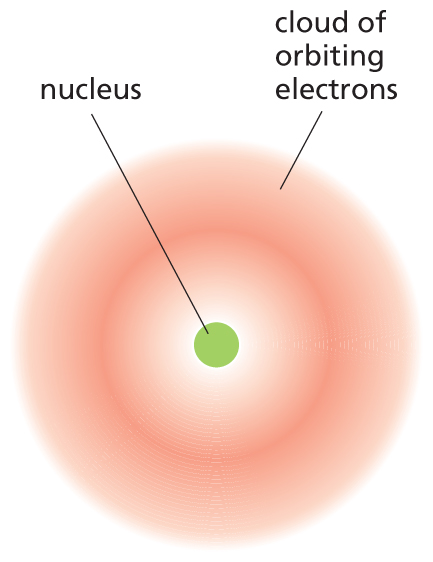
An illustration shows an atomic model. The model shows a spherical nucleus at the center, surrounded by a cloud of orbiting electrons.
Figure 2–1An atom consists of a nucleus surrounded by an electron cloud. The dense, positively charged nucleus contains nearly all of the atom’s mass. The much lighter and negatively charged electrons occupy space around the nucleus, as governed by the laws of quantum mechanics. The electrons are depicted as a continuous cloud, because there is no way of predicting exactly where an electron is at any given instant. The density of shading of the cloud is an indication of the probability that electrons will be found there.
The diameter of the electron cloud ranges from about 0.1 nm (for hydrogen) to about 0.4 nm (for atoms of high atomic number). The nucleus is very much smaller: about 5 × 10–6 nm for carbon, for example. If this diagram were drawn to scale, the nucleus would not be visible.
Each atom has at its center a dense, positively charged nucleus; this structure is surrounded at some distance by a cloud of negatively charged electrons, held in specific orbits by electrostatic attraction to the nucleus (Figure 2–1). The nucleus consists of two kinds of subatomic particles: protons, which are positively charged, and neutrons, which are electrically neutral. The atomic number of an element is determined by the number of protons present in the nucleus of its atoms. An atom of hydrogen, for example, has a nucleus composed of a single proton; with an atomic number of 1, hydrogen is the lightest element. An atom of carbon has six protons in its nucleus and an atomic number of 6 (Figure 2–2).
More information
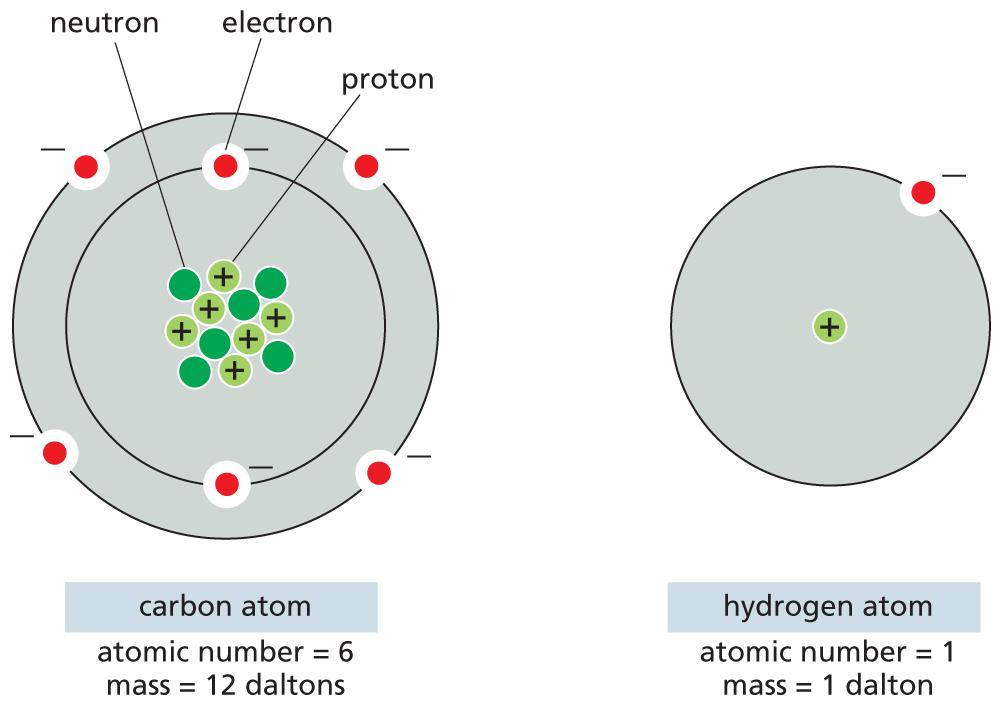
The atomic models of carbon and hydrogen are shown. Carbon atom: There are 6 neutrons, and 6 protons at the center of the atom. The atom has two orbitals; the inner orbital has 2 electrons and the outer orbital has 4 electrons. Atomic number equals 6, and the mass equals 12 daltons.
Hydrogen atom: The atom has one orbital with a proton at the center. A single electron orbits the nucleus. Atomic number equals 1 and the mass equals 1 dalton.
Figure 2–2The number of protons in an atom determines its atomic number. Schematic representations of an atom of carbon and an atom of hydrogen are shown. The nucleus of every atom except hydrogen consists of both positively charged protons and electrically neutral neutrons; the mass of an atom equals the number of protons plus neutrons. The number of electrons in an atom is equal to the number of protons, so that the atom has no net charge.
Isotopes of these elements would have the same number of protons, but additional neutrons (not shown). 14C, for example, has six protons but eight neutrons. Isotopes of hydrogen also exist: deuterium (2H) has a single neutron and tritium (3H) has two neutrons in addition to its proton.
In contrast to Figure 2–1, the electrons here are shown as individual particles. The concentric black circles represent in a highly schematic form the “orbits” (i.e., the different distributions) of the electrons. The neutrons, protons, and electrons are in reality minuscule in relation to the atom as a whole; their size is greatly exaggerated in these diagrams.
The electric charge carried by each proton is exactly equal and opposite to the charge carried by a single electron. Because the whole atom is electrically neutral, the number of positively charged protons contained in the nucleus will be balanced by an equal number of negatively charged electrons surrounding the nucleus; thus, the number of electrons in an atom, like the number of protons, matches its atomic number. All atoms of a given element have the same atomic number, and we will see shortly that it is this number that dictates each element’s chemical behavior.
Neutrons contribute to the structural stability of the nucleus: if there are too many or too few, the nucleus may disintegrate by radioactive decay. However, neutrons do not alter the chemical properties of the atom. Thus an element can exist in several physically distinguishable but chemically identical forms, called isotopes, each having a different number of neutrons but the same number of protons. Multiple isotopes of almost all the elements occur naturally, including some that are unstable—and thus radioactive. For example, while most carbon on Earth exists as carbon 12, a stable isotope with six protons and six neutrons, also present are small amounts of an unstable isotope, carbon 14, which has six protons and eight neutrons. Carbon 14 undergoes radioactive decay at a slow but steady rate, a property that allows archaeologists to estimate the age of organic material—a technique called “radiocarbon dating” or simply “carbon dating.”
Atoms are so small that it is hard to imagine how they can be detected and analyzed. An individual carbon atom is roughly 0.2 nm in diameter, so it would take about 5 million of them, laid out in a straight line, to span a millimeter. Similarly, the mass of an atom is so minuscule that chemists measure it using a special unit called a dalton, named after the English chemist who first described atoms as being the fundamental building blocks of matter. The major isotope of carbon, written as 12C, has a mass of 12 daltons; the unstable carbon isotope, 14C, has a mass of 14 daltons. Indeed, a dalton is defined as being one-twelfth the mass of a single 12C atom. Because electrons are so light, they contribute almost nothing to this mass. An atom’s mass therefore depends on the number of protons and neutrons in its nucleus, each of which has a mass of 1 dalton.
More information
A textbox has examples for moles and molar solutions. A mole represents a fixed amount of a substance. One mole contains exactly 6.022 times 10 to the 23rd particles of a substance. For an atom with a mass of X daltons, 1 mole of that atom will have a mass of X grams. The same is true for ions or molecules. A carbon atom has a mass of 12 daltons; 1 mole of carbon has a mass of 12 grams. A glucose molecules has a mass of 180 daltons; 1 mole of glucose has a mass of 180 grams. A sodium ion has a mass of 23 daltons and a chloride ion has a mass of 35 daltons; 1 mole of sodium chloride has a mass of 58 grams. A 1 molar solution has a concentration of 1 mole of the substance in 1 liter of solution. A 1 molar solution of glucose, for example, contains 180 grams per liter, and a one millimolar solution contains 180 milligrams per liter. The standard abbreviation for gram is a lower case g; the abbreviation for liter is an upper case L.
Figure 2–3Chemists measure out materials in moles. If two chemicals combine in a fixed proportion—for example, Na+ + Cl–⟶ NaCl—a chemist wishing to produce sodium chloride would, in theory, combine equal numbers of sodium and chloride ions. But how can someone count individual ions or atoms? To be able to mix chemicals in the correct proportions, scientists came up with the concept of a mole. A mole contains exactly 6.022 × 1023 particles (whether atoms, molecules, or ions) of a substance. For an atom with a mass of X daltons, 6.022 × 1023 of those atoms will have a mass of X grams. So, for example, 23 grams of Na+ will have the same number of ions as 35 grams of Cl–.
Determining how many atoms (or ions or molecules) are in a mole would take the hard work and ingenuity of numerous scientists, and in recognition of his contribution, this value has been designated Avogadro’s number. Some simple examples of moles and molar solutions are shown.
But chemists don’t typically work with individual atoms of carbon or hydrogen. They have therefore come up with a quantity that represents a defined amount of a substance that will allow them to work with that material in the lab. This quantity, called a mole, contains 6.022 × 1023 particles of a substance—where the particles can be atoms, molecules, or ions. This enormous number, called Avogadro’s number, allows us to relate individual atoms or molecules to everyday quantities, and it allows chemists to weigh out the amount of material needed for chemical reactions. Because of how a mole is defined, if an atom has a mass of X daltons, then one mole of that atom—6.022 × 1023 particles—will have a mass of X grams. Thus, 1 mole of hydrogen—that is, 6.022 × 1023 hydrogen atoms—will have a mass of 1 gram (Figure 2–3).
About 90 different elements occur in nature, each differing from the others in the number of protons and electrons in its atoms. Living things, however, are made of only a small selection of these elements, four of which—carbon (C), hydrogen (H), nitrogen (N), and oxygen (O)—constitute 96% of any organism’s mass. This composition differs markedly from that of the nonliving, inorganic environment on Earth (Figure 2–4), and it reflects the fact that a distinctive type of chemistry operates in biological systems.
More information
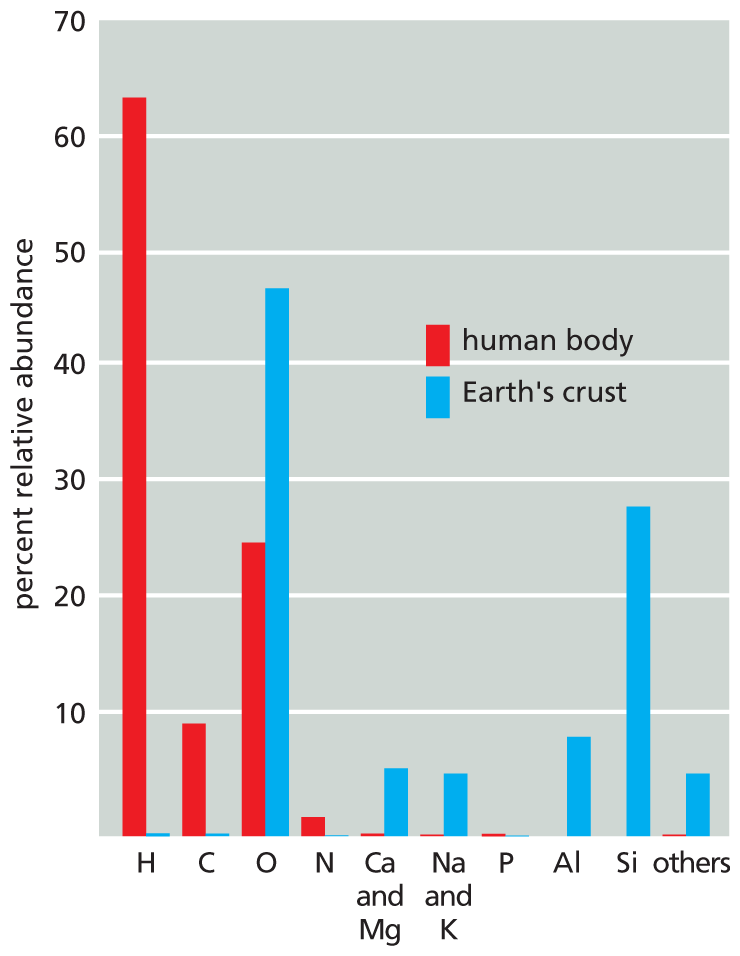
A bar graph depicts the abundance of various elements in the human body and the Earth’s crust. The horizontal axis shows the elements and the vertical axis shows percent relative abundance ranging from 10 to 70 in increments of 10. The data in the table reads as follows:
element-hydrogen, human body-63 percent, Earth’s crust-0.5 percent; element-carbon, human body-9 percent, Earth’s crust-0.5 percent; element-oxygen, human body-25 percent, Earth’s crust-47 percent; element-nitrogen, human body-2 percent, Earth’s crust-0.5 percent; element-carbon and magnesium, human body-0.5 percent, Earth’s crust-6 percent; element-sodium and potassium, human body-0.5 percent, Earth’s crust-5 percent; element-phosphorous, human body-0.5 percent, Earth’s crust-0.4 percent; element-aluminum, human body-0 percent, Earth’s crust-8 percent; element-silicon, human body-0 percent, Earth’s crust-28 percent; element-others, human body-0.5 percent, Earth’s crust-5 percent.
Figure 2–4The distribution of elements in the Earth’s crust differs radically from that in the human body. The abundance of each element is expressed here as a percentage of the total number of atoms present in a biological or geological sample (water included). Thus, for example, more than 60% of the atoms in the human body are hydrogen atoms, and nearly 30% of the atoms in the Earth’s crust are silicon atoms (Si). The relative abundance of elements is similar in all living things.
The Outermost Electrons Determine How Atoms Interact
To understand how atoms come together to form the molecules that make up living organisms, we have to pay special attention to each atom’s electrons. Protons and neutrons are welded tightly to one another in an atom’s nucleus, and they change partners only under extreme conditions—during radioactive decay, for example, or in the interior of the sun or a nuclear reactor. In living tissues, only the electrons of an atom undergo rearrangements. They form the accessible part of the atom and determine the chemical rules by which atoms combine to form molecules.
Electrons are in continuous motion around the nucleus, but motions on this submicroscopic scale obey laws that differ from those we are familiar with in everyday life. These laws dictate that electrons in an atom can exist only in certain discrete regions of movement—very roughly speaking, in distinct orbits. Each of these orbits, called an electron shell, can accommodate a limited number of electrons, based in part on its distance from the positively charged nucleus. The electrons closest on average to the nucleus are attracted most strongly to it and occupy the inner, most tightly bound shell. This innermost shell can hold a maximum of two electrons. The second shell is farther away from the nucleus, and can hold up to eight electrons. The third shell can also hold up to eight electrons, which are even less tightly bound. The fourth and fifth shells can hold 18 electrons each. Atoms with more than four shells are very rare in biological molecules.
QUESTION 2–1
A cup containing exactly 18 g, or 1 mole, of water was emptied into the Aegean Sea 3000 years ago. What are the chances that the same quantity of water, scooped today from the Pacific Ocean, would include at least one of these ancient water molecules? Assume that the original sample has become evenly distributed across the world’s oceans, which contain approximately 1.5 billion cubic kilometers (1.5 × 109 km3) of water. For the purposes of this exercise, assume that for both the original water and for seawater, the density is 1 g/cm3.
The arrangement of electrons in an atom is most stable when all the electrons are in the most tightly bound states that are possible for them—that is, when they occupy the innermost shells, closest to the nucleus. Therefore, with certain exceptions in the larger atoms, the electrons of an atom fill the shells in order—the first before the second, the second before the third, and so on. An atom whose outermost shell is entirely filled with electrons is especially stable and therefore chemically unreactive. Examples are helium with 2 electrons (atomic number 2), neon with 2 + 8 electrons (atomic number 10), and argon with 2 + 8 + 8 electrons (atomic number 18); these are all inert gases. Hydrogen, by contrast, has only one electron; because its outermost shell is only half-filled, hydrogen is therefore highly reactive. The atoms found in living organisms all have outermost shells that are incompletely filled, which allows them to react with one another to form molecules (Figure 2–5).
More information
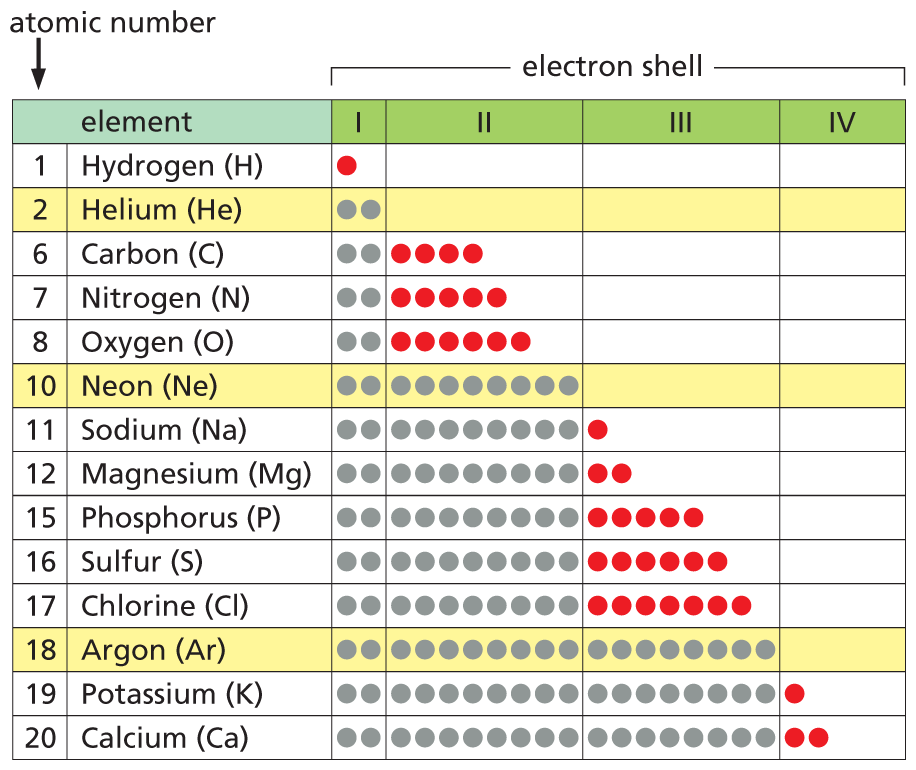
A table has arranged various elements by increasing atomic number and the degree of completion of their electron shells. For each element, there are four columns representing four levels of electron shells. Hydrogen is listed first at atomic number 1. The first shell is incompletely filled with a single electron. Helium, at atomic number 2, completely fills the first shell with 2 electrons. Carbon, at atomic number 6, completely fills the first shell with 2 electrons and incompletely fills the second shell with 4 electrons. Nitrogen, at atomic number 7, completely fills the first shell with 2 electrons and incompletely fills the second shell with 5 electrons. Oxygen, at atomic number 8, completely fills the first shell with 2 electrons, and incompletely fills the second shell with 6 electrons. Neon, at atomic number 10, completely fills both the first and second shells, with 2 and 8 electrons respectively. Sodium, at atomic number 11, completely fills shells 1 and 2, and incompletely fills shell 3 with 1 electron. Magnesium, at atomic number 12, completely fills the first 2 shells and incompletely fills the third shell with 2 electrons. Phosphorus, at atomic number 15, completely fills the first 2 shells and incompletely fills the third shell with 5 electrons. Sulfur, at atomic number 16, completely fills the first 2 shells and incompletely fills the third shell with 6 electrons. Chlorine, at atomic number 17, completely fills the first 2 shells and incompletely fills the third shell with 7 electrons. Argon, at atomic number 18, completely fills the first 3 shells with 2, 8, and 8 electrons respectively. Potassium, at atomic number 19, completely fills the first 3 shells, and incompletely fills the fourth shell with 1 electron. Calcium, at atomic number 20, completely fills the first 3 shells, and incompletely fills the fourth shell with 2 electrons. The inert gases in the table are Helium, Neon, and Argon. These elements do not have incomplete electron shells.
Figure 2–5An element’s chemical reactivity depends on the degree to which its outermost electron shell is filled. All of the elements commonly found in living organisms have outermost shells that are not completely filled. The electrons in these incomplete shells (here shown in red) can participate in chemical reactions with other atoms. Inert gases (yellow), in contrast, have completely filled outermost shells (gray) and are thus chemically unreactive.
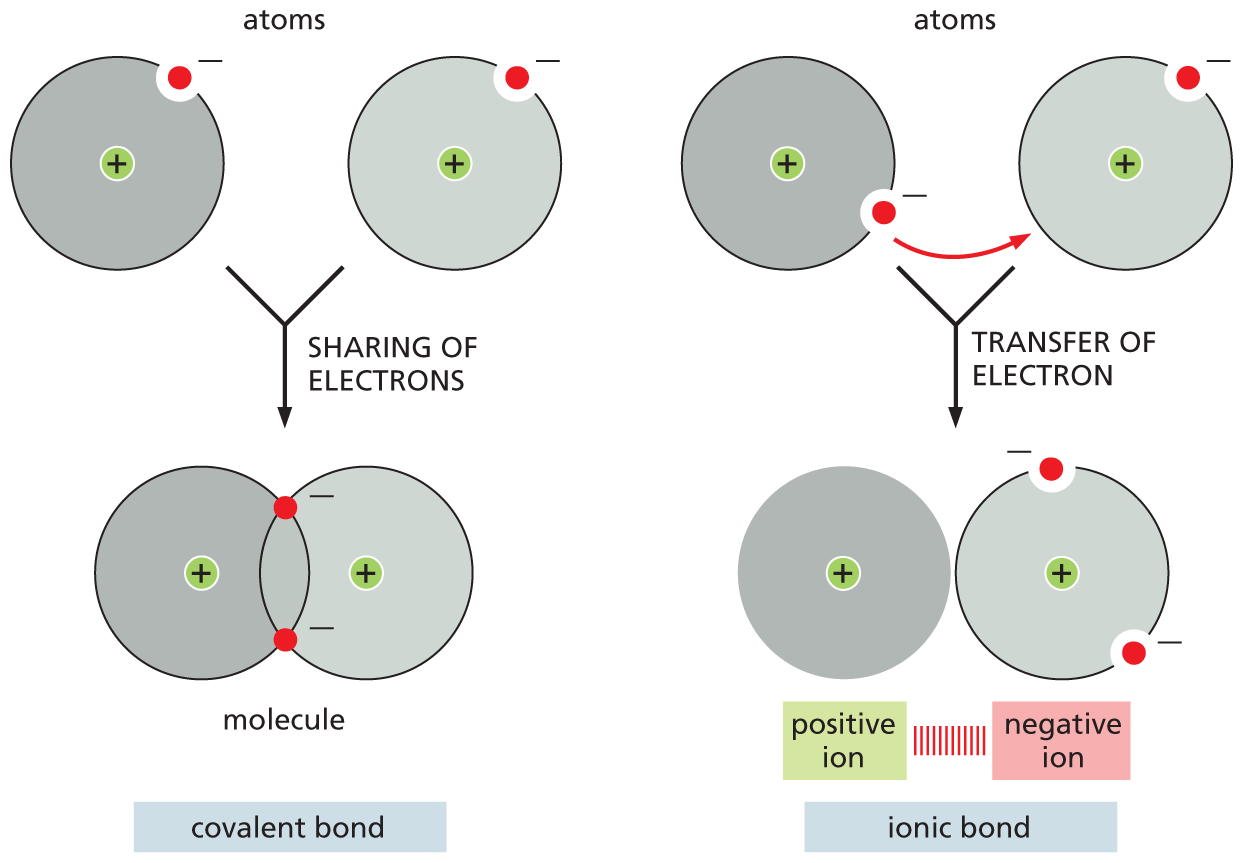
Because an incompletely filled electron shell is less stable than one that is completely filled, atoms with incomplete outer shells have a strong tendency to interact with other atoms so as to either gain or lose enough electrons to fill their outermost shell. This electron exchange can be achieved either by transferring electrons from one atom to another or by sharing electrons between two atoms. These two strategies produce two types of chemical bonds that can bind atoms strongly to one another: an ionic bond is formed when electrons are donated by one atom to another, whereas a covalent bond is formed when two atoms share a pair of electrons (Figure 2–6).
More information
Two illustrations depict the formation of a covalent bond and an ionic bond. Covalent bond: two atoms each with a proton and an electron share their electrons to form a molecule. The molecule is depicted as two overlapping atoms; the overlapping region has two electrons that are shared between the two atoms.
Ionic bond: two atoms each with a proton and an electron are shown; an electron from the first atom is transferred to the second atom; the first atom forms a positive ion (no electrons) and the second atom forms a negative ion (two electrons).
Figure 2–6Atoms can attain a more stable arrangement of electrons in their outermost shell by interacting with one another. A covalent bond is formed when electrons are shared between atoms. An ionic bond is formed when electrons are transferred from one atom to the other, and the resulting ions are held together by electrostatic attraction. The two cases shown represent extremes; often, covalent bonds form with a partial transfer (unequal sharing) of electrons, resulting in a polar covalent bond, as we discuss shortly.
An H atom, which needs only one more electron to fill its only shell, generally acquires this electron by sharing—forming one covalent bond with another atom. The other most common elements in living cells—C, N, and O, which have an incomplete second shell, and P and S, which have an incomplete third shell (see Figure 2–5)—also tend to share electrons; these elements fill their outer shells by forming several covalent bonds. The number of electrons an atom must acquire or lose (either by sharing or by transfer) to attain a filled outer shell determines the number of bonds that the atom can make.
QUESTION 2–2
A carbon atom contains six protons and six neutrons.
What are its atomic number and mass?
How many electrons does it have?
How many additional electrons must it add to fill its outermost shell? How does this affect carbon’s chemical behavior?
Carbon with a mass of 14 daltons is radioactive. How does it differ in structure from nonradioactive carbon? How does this difference affect its chemical behavior?
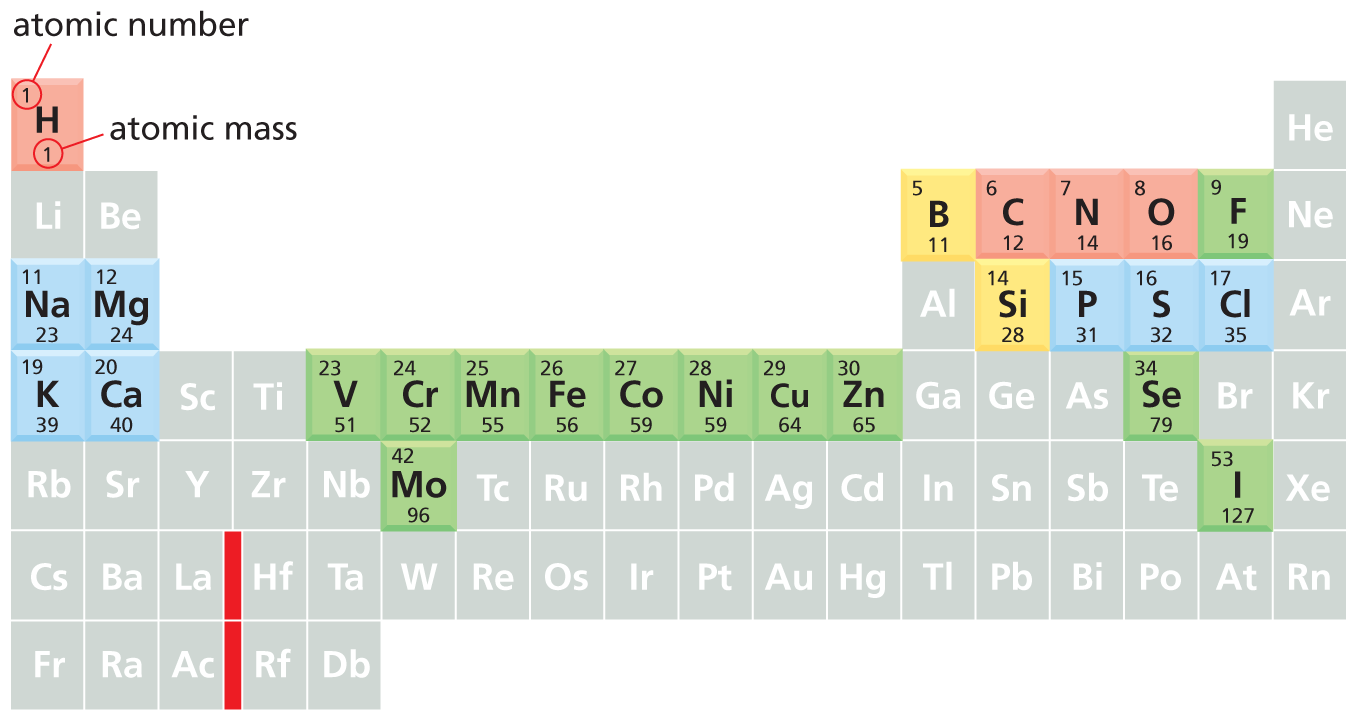
Because the state of the outer electron shell determines the chemical properties of an element, when the elements are listed in order of their atomic number we see a periodic recurrence of elements that have similar properties. For example, an element with an incomplete second shell containing one electron will behave in a similar way as an element that has filled its second shell and has an incomplete third shell containing one electron. And, as we have just seen, the atoms that have full outer shells, the noble gases, are chemically inert. This chemical consistency gives rise to the famous periodic table of the elements, outlined in Figure 2–7, in which we have highlighted the elements found in living organisms.
More information
An illustration shows a periodic table where elements that have similar properties are highlighted. The periodic table has seven rows and eighteen columns. The different groupings are highlighted in different colors as follows:
Pink: H (atomic number-1, atomic mass-1), C (atomic number-6, atomic mass-12), N (atomic number-7, atomic mass-14), O (atomic number-8, atomic mass-16), F (atomic number-9, atomic mass-19).
Blue: N a (atomic number-11, atomic mass-23), M g (atomic number-12, atomic mass-24), K (atomic number-19, atomic mass-39), C a (atomic number-20, atomic mass-40), P (atomic number-15, atomic mass-31), S (atomic number-16, atomic mass-32), C l (atomic number-17, atomic mass-35).
Green: V (atomic number-23, atomic mass-51), C r (atomic number-24 , atomic mass-52 ), M n (atomic number- 25 , atomic mass-55), F e (atomic number-26 , atomic mass-56 ), C o (atomic number-27 , atomic mass-59), N i (atomic number-28 , atomic mass-59), C u (atomic number-29 , atomic mass-64 ), Z n (atomic number-30 , atomic mass-65), M o (atomic number-42 , atomic mass-96 ), F (atomic number-9 , atomic mass-19), S e (atomic number-34 , atomic mass-79 ), I (atomic number-53 , atomic mass-127 ).
Yellow: B (atomic number-5 , atomic mass-11 ), S i (atomic number-14 , atomic mass-28 ).
The remaining elements are highlighted in gray: L i, R b, C s, F r, B e, S r, B a, R a, S c, Y, L a, A c, T i, Z r, H f, R f, N b, T a, D b, W, T c, R e, R u, O s, R h, I r, P d, P t, A g, A u, C d, H g, A l, G a, l n, T l, G e, S n, P b, A s, S b, B i, T e, P o, A t, B r, H e, N e, A r, K r, X e, and R n.
Figure 2–7When ordered by their atomic number into the periodic table, the elements fall into vertical columns in which the atoms have similar properties. Atoms in the same vertical column must gain or lose the same number of electrons to attain a filled outer shell (see Figure 2–5); they therefore behave similarly when forming bonds with other atoms. Thus, for example, both magnesium (Mg) and calcium (Ca) tend to give away the two electrons in their outer shells to form ionic bonds with atoms such as chlorine (Cl), which need extra electrons to complete their outer shells.
The chemistry of life is dominated by lighter elements. The four elements highlighted in red constitute 99% of the total number of atoms present in the human body and about 96% of our total mass. An additional seven elements, highlighted in blue, together represent about 0.9% of our total number of atoms. Other elements, shown in green, are required in trace amounts by humans. It remains unclear whether those elements shown in yellow are essential in humans or not.
The masses listed here are those of the most common isotope of each element. The vertical red line represents a break in the periodic table where a group of large atoms with similar chemical properties has been removed.
Covalent Bonds Form by the Sharing of Electrons
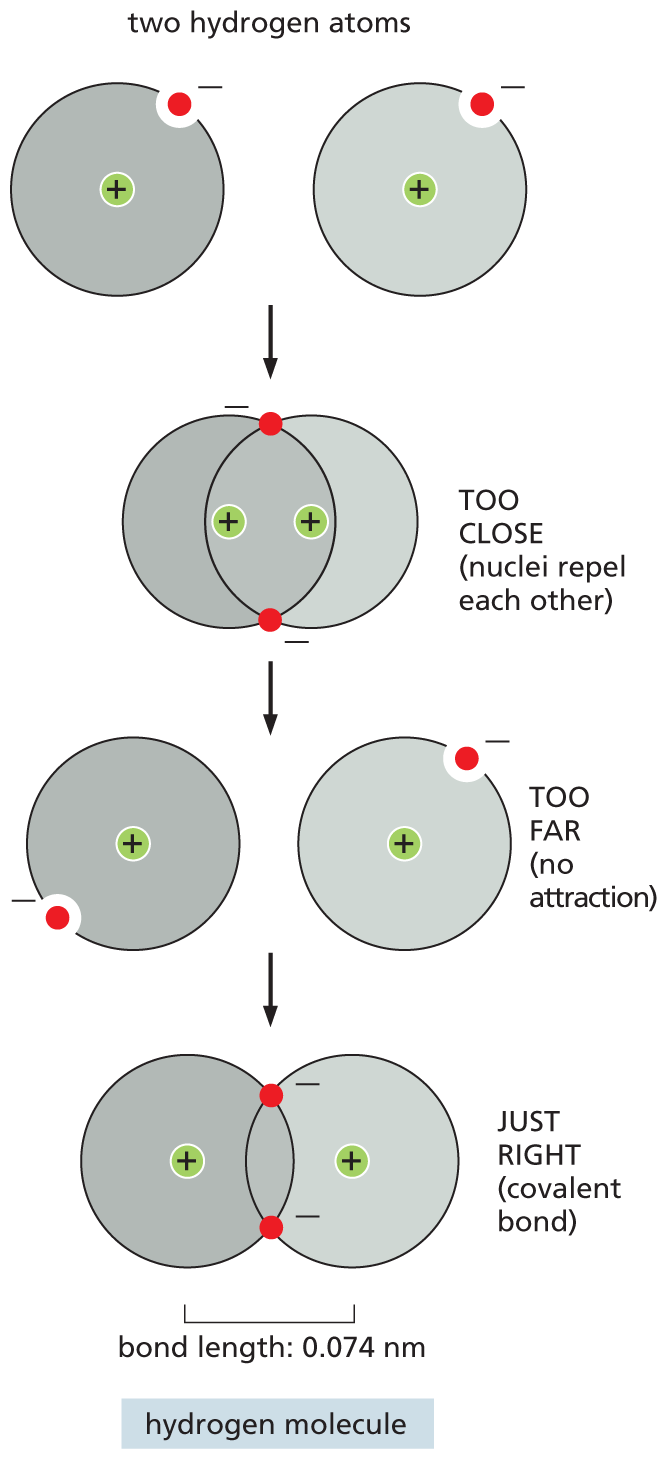
All of the characteristics of a cell depend on the molecules it contains. A molecule is a cluster of atoms held together by covalent bonds, in which electrons are shared rather than transferred between atoms. The shared electrons complete the outer shells of the interacting atoms. In the simplest possible molecule—a molecule of hydrogen gas (H2)—two H atoms, each with a single electron, share their electrons, thus filling their outermost shells. The shared electrons form a cloud of negative charge that is densest between the two positively charged nuclei. This electron density helps to hold the nuclei together by opposing the mutual repulsion between the positive charges of the nuclei, which would otherwise force them apart. The attractive and repulsive forces are precisely in balance when these nuclei are separated by a characteristic distance, called the bond length (Figure 2–8).
More information
An illustration shows incorrect and correct bonding of two hydrogen atoms in a covalent bond. A covalent bond is successful when the nuclei of the two atoms are at a distance of 0.074 nanometers from each other. Two hydrogen atoms covalently bound are referred to as a hydrogen molecule, H 2. Nuclei that are closer than this distance repel each other and will not form a covalent bond. Nuclei that are farther than this distance will not be able to share electrons effectively and will not attract each other.
Figure 2–8A hydrogen molecule is held together by a covalent bond of a particular length. Each hydrogen atom in isolation has a single electron, which means that its first (and only) electron shell is incompletely filled. By coming together to form a hydrogen molecule (H2, or hydrogen gas), the two atoms are able to share their electrons, so that each obtains a completely filled first shell, with the shared electrons adopting modified orbits around the two nuclei. The covalent bond between the two atoms has a defined length—0.074 nm, which is the distance between the two nuclei. If the atoms were closer together, the positively charged nuclei would repel each other; if they were farther apart, they would not be able to share electrons as effectively.
Whereas an H atom can form only a single covalent bond, the other common atoms that form covalent bonds in cells—O, N, S, and P, as well as the all-important C—can form more than one. The outermost shells of these atoms, as we have seen, can accommodate up to eight electrons, and they form covalent bonds with as many other atoms as necessary to reach this number. Oxygen, with six electrons in its outer shell, is most stable when it acquires two extra electrons by sharing with other atoms, and it therefore forms up to two covalent bonds. Nitrogen, with five outer electrons, forms a maximum of three covalent bonds, while carbon, with four outer electrons, forms up to four covalent bonds—thus sharing four pairs of electrons (see Figure 2–5).
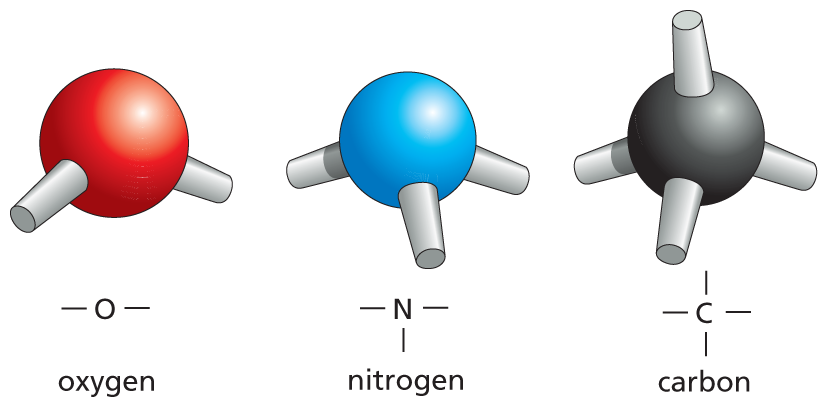
When one atom forms covalent bonds with several others, these multiple bonds have definite orientations in space relative to one another, reflecting the orientations of the orbits of the shared electrons. Covalent bonds between multiple atoms are therefore characterized by specific bond angles, as well as by specific bond lengths and bond energies (Figure 2–9). The four covalent bonds that can form around a carbon atom, for example, are arranged as if pointing to the four corners of a regular tetrahedron. The precise orientation of the covalent bonds around carbon dictates the three-dimensional geometry of all organic molecules.
More information
Oxygen: a red sphere with two single sticks. Representation: single bond O single bond.
Nitrogen: a blue sphere with three single sticks. Representation: a central N, attached to three single bonds.
Carbon: a black sphere with four single sticks. Representation: a central C, attached to four single bonds.
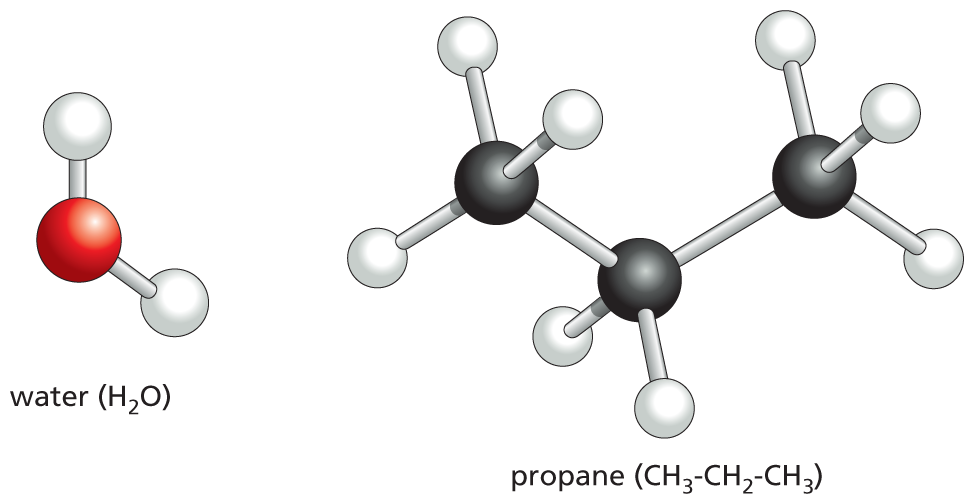
(A)More information
Water (H subscript 2 O): a red sphere single bonded to two white spheres.
Propane (C H subscript 3 C H subscript 2 C H subscript 3): a chain of three black spheres; the terminal black spheres are each single bonded to three white spheres; the middle black sphere is single bonded to two white spheres.
(B)Figure 2–9Covalent bonds are characterized by particular geometries. (A) The spatial arrangement of the covalent bonds that can be formed by oxygen, nitrogen, and carbon. (B) Molecules formed from these atoms therefore have precise three-dimensional structures defined by the bond angles and bond lengths for each covalent linkage. A water molecule, for example, forms a “V” shape with an angle of about 109°.
In these ball-and-stick models, the different colored balls represent different atoms, and the sticks represent the covalent bonds. The colors traditionally used to represent the different atoms—black (or sometimes dark gray) for carbon, white for hydrogen, blue for nitrogen, and red for oxygen—were established by the chemist August Wilhelm Hofmann in 1865, when he used a set of colored croquet balls to build molecular models for a public lecture on “the combining power of atoms.”
Some Covalent Bonds Involve More Than One Electron Pair
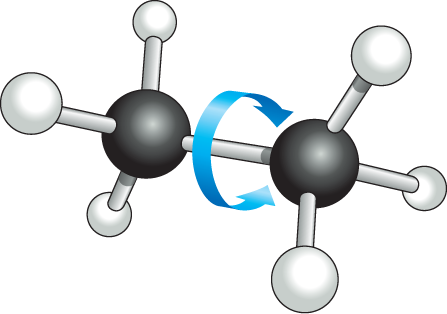
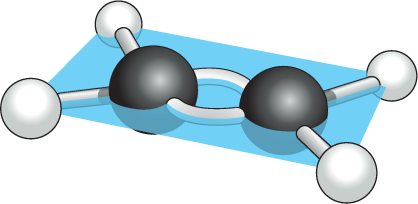
Most covalent bonds involve the sharing of two electrons, one donated by each participating atom; these are called single bonds. Some covalent bonds, however, involve the sharing of more than one pair of electrons. In a double bond, for example, four electrons are shared, two coming from each participating atom. Double bonds are shorter and stronger than single bonds and have a characteristic effect on the geometry of molecules containing them. A single covalent bond between two atoms generally allows the rotation of one part of a molecule relative to the other around the bond axis. A double bond prevents such rotation, producing a more rigid and less flexible arrangement of atoms (Figure 2–10). This restriction has a major influence on the three-dimensional shape of many macromolecules.
More information
The ball and stick model of ethane consists of 2 carbon atoms bound to each other at 180 degrees with a single bond. Each of the carbon atoms are also bound to 3 hydrogen atoms in a tetrahedral arrangement. An arrow curving around the carbon to carbon bond indicates that the C H subscript 3 groups can rotate around the bond axis.
(A) ethaneMore information
The ball and stick model of ethene consists of 2 carbon atoms bound to each other at 180 degrees with a double bond. Each carbon atom is also bound to 2 hydrogen atoms at bent angles. All atoms in the ethene model are in the same plane.
(B) ethaneFigure 2–10Carbon–carbon double bonds are shorter and more rigid than carbon–carbon single bonds. (A) The ethane molecule, with a single covalent bond between its two carbon atoms (black), shows the tetrahedral arrangement of the three single covalent bonds each carbon atom shares with its three attached H atoms. The CH3 groups, joined by a covalent C–C bond, can rotate relative to one another around the bond axis. (B) The double bond between the two carbon atoms in a molecule of ethene (ethylene) alters the bond geometry of the carbon atoms and brings all the atoms into the same plane; the double bond prevents the rotation of one CH2 group relative to the other.
Some molecules contain atoms that share electrons in a way that produces bonds that are intermediate in character between single and double bonds. The highly stable benzene molecule, for example, is made up of a ring of six carbon atoms in which the bonding electrons are evenly distributed, although the arrangement is sometimes depicted as an alternating sequence of single and double bonds. Panel 2–1 (pp. 70–71) reviews the covalent bonds commonly encountered in biological molecules.
Electrons in Covalent Bonds Are Often Shared Unequally
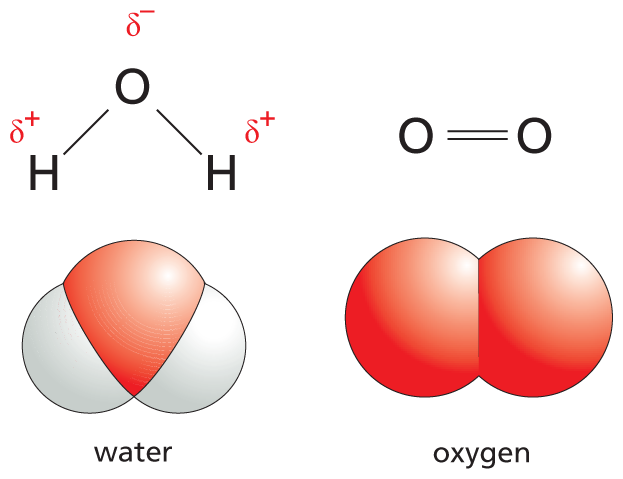
When the atoms joined by a single covalent bond belong to different elements, the two atoms usually attract the shared electrons to different degrees. Covalent bonds in which the electrons are shared unequally in this way are known as polar covalent bonds. A polar structure (in the electrical sense) is one in which a positive charge is concentrated toward one atom in the molecule (the positive pole) and a negative charge is concentrated toward another atom (the negative pole). The tendency of an atom to attract electrons is called its electronegativity, a property that was first described by the chemist Linus Pauling.
Knowing the electronegativity of atoms allows one to predict the nature of the bonds that will form between them. For example, when atoms with different electronegativities are covalently linked, their bonds will be polarized. Among the atoms typically found in biological molecules, oxygen and nitrogen (with electronegativities of 3.4 and 3.0, respectively) attract electrons relatively strongly, whereas an H atom (with an electronegativity of 2.1) attracts electrons relatively weakly. Thus the covalent bonds between O and H (O–H) and between N and H (N–H) are polar (Figure 2–11). An atom of C, by contrast, has an electronegativity (2.6) that is closer to that of hydrogen. Because these atoms attract electrons more equally, the bond between carbon and hydrogen, C–H, is relatively nonpolar.
More information
An illustration shows skeletal structures and space filling models of water and oxygen. The skeletal structure of the water molecule shows a central oxygen atom single bonded to 2 hydrogen atoms. The oxygen atom has a partial negative charge and the hydrogen atoms have partial positive charges. The space filling model of the water molecule shows the central oxygen atom with 2 hydrogen atoms heavily overlapping the oxygen atom at a bent angle.
The skeletal structure of the oxygen molecule shows 2 double bonded oxygen atoms. The space filling model shows 2 oxygen atoms fused together.
Figure 2–11The polarity of a covalent bond depends on the relative electronegativities of the participating atoms. Shown here are the electron distributions for the polar covalent bonds in a molecule of water (H2O) and the nonpolar covalent bonds in a molecule of oxygen (O2). In the O2 molecule, the electrons are evenly distributed, whereas in H2O, electrons are more strongly attracted to the oxygen nucleus than to the H nucleus, producing partial negative (δ–) and partial positive (δ+) charges.
Covalent Bonds Are Strong Enough to Survive the Conditions Inside Cells
The covalent bond between two atoms has a characteristic length that depends on the atoms involved. A further crucial property of any chemical bond is its strength. Bond strength is measured by the amount of energy that must be supplied to break the bond, usually expressed in units of either kilocalories per mole (kcal/mole) or kilojoules per mole (kJ/mole). A kilocalorie is the amount of energy needed to raise the temperature of 1 liter of water by 1°C. Thus, if 1 kilocalorie of energy must be supplied to break 6 × 1023 bonds of a specific type (that is, 1 mole of these bonds), then the strength of that bond is 1 kcal/mole. One kilocalorie is equal to about 4.2 kJ, which is the unit of energy universally employed by physical scientists and, increasingly, by cell biologists as well.
To get an idea of what bond strengths mean, it is helpful to compare them with the average energies of the impacts that molecules continually undergo owing to collisions with other molecules in their environment—their thermal, or heat, energy. Typical covalent bonds are stronger than these thermal energies by a factor of 100, so they are resistant to being pulled apart by thermal motions. In living organisms, covalent bonds are normally broken only during specific chemical reactions that are carefully controlled by highly specialized protein catalysts called enzymes.
Ionic Bonds Form by the Gain and Loss of Electrons
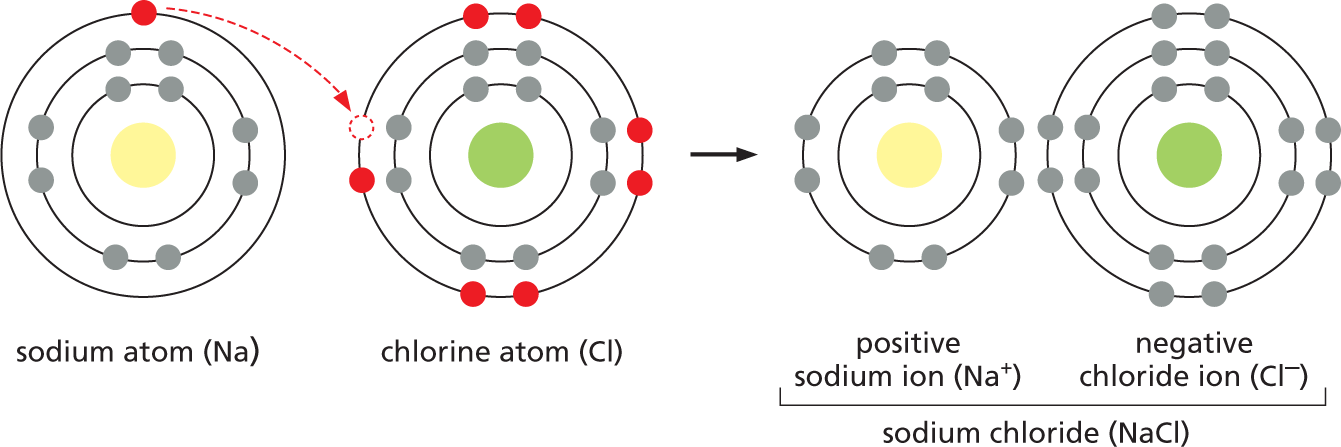
In some substances, the participating atoms are so different in electronegativity that their electrons are not shared at all—they are transferred completely to the more electronegative partner. Atoms that engage in these electron transfers are generally those that can attain a completely filled outer shell most easily by donating electrons to—or accepting electrons from—another atom, rather than by sharing them. For example, a sodium (Na) atom can achieve a filled outer shell by giving up the single electron in its third shell, whereas a chlorine (Cl) atom can complete its outer shell by gaining just one electron (see Figure 2–5). As a consequence, if a sodium atom encounters a chlorine atom, an electron can jump from the Na to the Cl, leaving both atoms with filled outer shells. The offspring of this marriage between sodium, a soft and intensely reactive metal, and chlorine, a toxic green gas, is table salt (NaCl).
QUESTION 2–3
Discuss whether the following statement is correct: “An ionic bond can, in principle, be thought of as a very polar covalent bond. Polar covalent bonds, then, fall somewhere between ionic bonds at one end of the spectrum and nonpolar covalent bonds at the other end.”
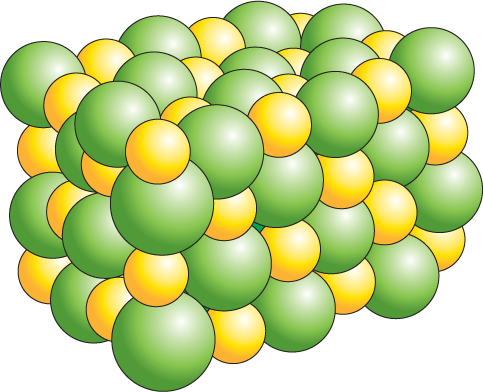
When an electron jumps from Na to Cl, both atoms become electrically charged ions. The Na atom that lost an electron now has one less electron than it has protons in its nucleus; it therefore has a net single positive charge (Na+). The Cl atom that gained an electron now has one more electron than it has protons and has a net single negative charge (Cl–). Because of their opposite charges, the Na+ and Cl– ions are attracted to each other; these electrostatic attractions allow Na+ and Cl– to form an ionic bond (Figure 2–12A). Ions held together solely by ionic bonds are generally called salts rather than molecules. A crystal of NaCl contains astronomical numbers of Na+ and Cl– ions packed together in a precise, three-dimensional array with their opposite charges exactly balanced: a crystal only 1 mm across contains about 2 × 1019 ions of each type (Figure 2–12B and C). Unlike our drawing of an electrostatic attraction between a single pair of ions in Figure 2–12A, each positively charged Na+ ion in a crystal of salt will bond with all of the negatively charged Cl– ions that surround it—and vice versa; it is this network of attractions that holds the crystal together.
More information
An illustration shows atomic models of sodium, chlorine, and sodium chloride. The atomic model of a sodium atom, N a , has a central nucleus surrounded by 3 orbitals. The inner orbital has 2 electrons, the middle orbital has 8 electrons, and the outer orbital has 1 electron. The inner and middle orbitals are completely filled. The outer orbital is incompletely filled. The atomic model of a chlorine atom, C l , shows a central nucleus surrounded by 3 orbitals. The inner orbital has 2 electrons, the middle orbital has 8 electrons, and the outer orbital has 7 electrons. The inner and middle orbitals are completely filled. The outer orbital is incompletely filled. When N a reacts with C l, 1 electron from the sodium atom moves to the outer orbital of the chlorine atom and forms sodium chloride, N a C l. The sodium atom gains a positive charge and is now referred to as a sodium ion, N a superscript plus. The sodium ion has two orbitals that are completely filled. The chlorine atom gains a negative charge and is now referred to as a chloride ion, C l superscript minus. The chloride ion has three orbitals that are completely filled. The outer orbital of the chloride ion has a total of 8 electrons.
(A)More information
An illustration shows a space filling model of sodium chloride, an array of tightly packed ions of sodium and chloride. The packing of the ions loosely forms a rectangular prism.
(B)More information
A micrograph shows crystals of sodium chloride, approximately 0.5 millimeters in diameter.
Figure 2–12Sodium chloride is held together by ionic bonds. (A) An atom of sodium (Na) reacts with an atom of chlorine (Cl). Electrons of each atom are shown in their different shells; electrons in the chemically reactive (incompletely filled) outermost shells are shown in red. The reaction takes place with the transfer of a single electron from sodium to chlorine, forming two electrically charged atoms, or ions, each with complete sets of electrons in their outermost shells. The two ions have opposite charge and are held together by electrostatic attraction. (B) The product of the reaction between sodium and chlorine, crystalline sodium chloride, contains sodium and chloride ions packed closely together in a regular array in which the charges are exactly balanced. (C) Photograph of crystals of sodium chloride.
Electrically charged ions are also strongly attracted to polar water molecules. Therefore, many salts (including NaCl) are highly soluble in water, where they dissociate into individual ions (such as Na+ and Cl–), each surrounded by a group of water molecules. Positive ions are called cations and negative ions are called anions. Small inorganic ions such as Na+, Cl–, K+, and Ca2+ play important parts in many biological processes, including the electrical activity of nerve cells, as we discuss in Chapter 12.
Because of the favorable interaction between ions and water, in cells ionic bonds are 10–100 times weaker than the covalent bonds that hold atoms together in molecules. Nevertheless, these weak interactions play an important role in the chemistry of living things, as we discuss shortly.
Hydrogen Bonds Are Important Noncovalent Bonds for Many Biological Molecules
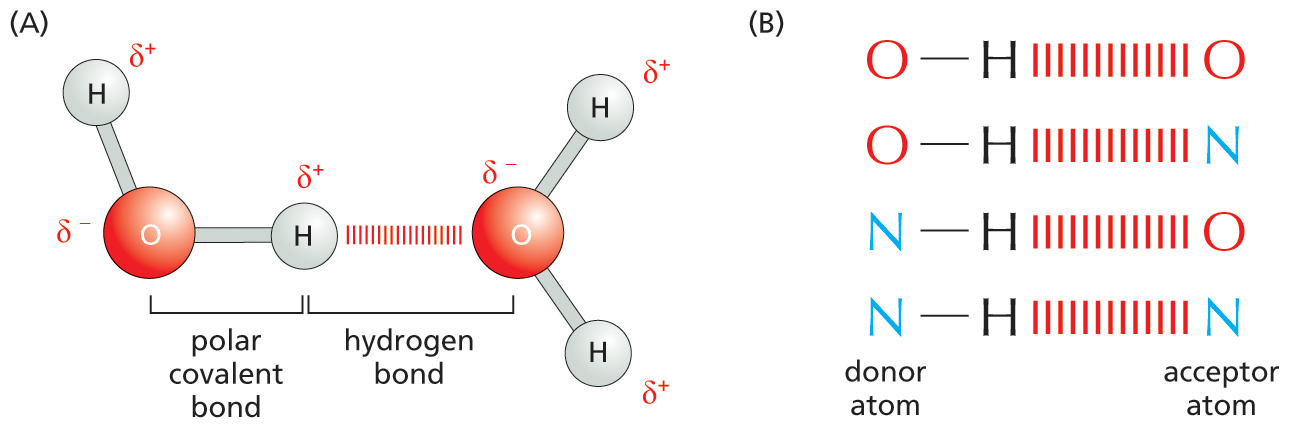
Water accounts for about 70% of a cell’s mass, and most intracellular reactions occur in an aqueous environment. Thus the properties of water have put a permanent stamp on the chemistry of living things. In each molecule of water (H2O), the two covalent H–O bonds are highly polar because the O is strongly attractive for electrons whereas the H is only weakly attractive. Consequently, in each water molecule, there is a preponderance of positive charge on the two H atoms and negative charge on the O. When a positively charged region of one water molecule (that is, one of its H atoms) comes close to a negatively charged region (that is, the O) of a second water molecule, the electrical attraction between them can establish a weak bond called a hydrogen bond (Figure 2–13A).
More information
A two-part illustration depicts hydrogen bonding between water molecules, and hydrogen bonding among other polar molecules.
Figure 2–13Noncovalent hydrogen bonds form between water molecules and between many other polar molecules. (A) A hydrogen bond forms between two water molecules. The slight positive charge associated with the hydrogen atom is electrically attracted to the slight negative charge of the nearby oxygen atom. (B) In cells, hydrogen bonds commonly form between molecules that contain an oxygen or nitrogen. The atom bearing the hydrogen is considered the H-bond donor and the atom that interacts with the hydrogen is the H-bond acceptor.
These bonds are much weaker than covalent bonds and are easily broken by random thermal motions. Thus each bond lasts only an exceedingly short time. But the combined effect of many weak bonds is far from trivial. Each water molecule can form hydrogen bonds through its two H atoms to two other water molecules, producing a network in which hydrogen bonds are being continually broken and formed. It is because of these interlocking hydrogen bonds that water at room temperature is a liquid—with a high boiling point and high surface tension—and not a gas. Without hydrogen bonds, life as we know it could not exist.
QUESTION 2–4
True or false? “When NaCl is dissolved in water, the water molecules closest to the ions will tend to preferentially orient themselves so that their oxygen atoms face the sodium ions and face away from the chloride ions.” Explain your answer.
Hydrogen bonds are not limited to water. In general, a hydrogen bond can form whenever a positively charged H atom held in one molecule by a polar covalent linkage comes close to a negatively charged atom—typically an oxygen or a nitrogen—belonging to another molecule (Figure 2–13B). Hydrogen bonds can also occur between different parts of a single large molecule, where they often help the molecule fold into a particular shape.
Like molecules (or salts) that carry positive or negative charges, substances that contain polar bonds and can form hydrogen bonds also mix well with water. Such substances are termed hydrophilic, meaning that they are “water-loving.” A large proportion of the molecules in the aqueous environment of a cell fall into this category, including sugars, DNA, RNA, and a majority of proteins. Hydrophobic (“water-fearing”) molecules, by contrast, are uncharged and form few or no hydrogen bonds, and they do not dissolve in water. These and other properties of water are reviewed in Panel 2–2 (pp. 72–73).
Four Types of Weak Interactions Help Bring Molecules Together in Cells
Much of biology depends on specific but transient interactions between one molecule and another. These associations are mediated by noncovalent bonds, such as the hydrogen bonds just discussed. Although these noncovalent bonds are individually quite weak, their energies can sum to create an effective force between two molecules.
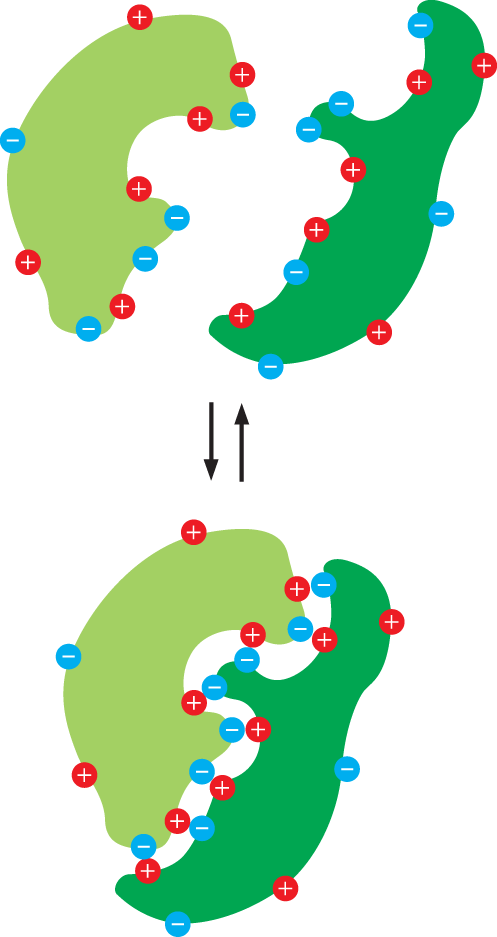
The ionic bonds that hold together the Na+ and Cl– ions in a salt crystal (see Figure 2–12) represent another form of noncovalent bond called an electrostatic attraction. Electrostatic attractions are strongest when the atoms involved are fully charged, as are Na+ and Cl– ions. But a weaker electrostatic attraction can occur between molecules that contain polar covalent bonds (see Figure 2–11). Like hydrogen bonds, electrostatic attractions are extremely important in biology. For example, any large molecule with many polar groups will have a pattern of partial positive and negative charges on its surface. When such a molecule encounters a second molecule with a complementary set of charges, the two will be drawn to each other by electrostatic attraction. Even though water greatly reduces the strength of these attractions in most biological settings, the large number of weak noncovalent bonds that form on the surfaces of large molecules can nevertheless promote strong and specific binding (Figure 2–14).
More information
An illustration depicts reversible binding between two protein molecules with positive and negative ions on their surfaces. The proteins are first shown separated. In the next step, the surfaces of the protein molecules come in contact with each other and the opposite charges attract, binding the protein molecules together. This molecule can again dissociate to form two separate protein molecules.
Figure 2–14A large molecule, such as a protein, can bind to another protein through noncovalent bonds on the surface of each molecule. In the aqueous environment of a cell, many individual weak interactions could cause the two proteins to recognize each other specifically and form a tight complex. Shown here is a set of electrostatic attractions between complementary positive and negative charges. (Dynamic Figure) A large molecule such as a protein will display a specific pattern of positive and negative charges on its surface. When two proteins encounter each other, there may be a number of weaker interactions before an alignment of complementary charges is recognized that facilitates strong and specific binding.
A third type of noncovalent bond, called a van der Waals attraction, comes into play when any two atoms approach each other closely. These nonspecific interactions spring from fluctuations in the distribution of electrons in every atom, which can generate a transient attraction when the atoms are in very close proximity. These weak attractions occur in all types of molecules, even those that are nonpolar and cannot form ionic or hydrogen bonds. The relative lengths and strengths of these three types of noncovalent bonds are compared to the length and strength of covalent bonds in Table 2–1.
TABLE 2–1 LENGTH AND STRENGTH*OF SOME CHEMICAL BONDS
The fourth effect that often brings molecules together is not, strictly speaking, a bond at all. In an aqueous environment, a hydrophobic force is generated by a pushing of nonpolar surfaces out of the hydrogen-bonded water network, where they would otherwise physically interfere with the highly favorable interactions between water molecules. Hydrophobic forces play an important part in promoting molecular interactions—in particular, in building cell membranes, which are constructed largely from lipid molecules with long hydrocarbon tails. In these molecules, the H atoms are covalently linked to C atoms by nonpolar bonds (see Panel 2–1, pp. 70–71). Because H atoms bound to carbon have almost no net positive charge, they cannot form effective hydrogen bonds to other molecules, including water. When pushed out of the network of water molecules inside cells, some lipids self-associate to form thin, sheetlike membrane barriers that keep the aqueous interior of the cell separate from the surrounding aqueous environment, a topic we return to later in the chapter.
All four types of weak chemical interactions important in biology are reviewed in Panel 2–3 (pp. 74–75).
Some Polar Molecules Form Acids and Bases in Water
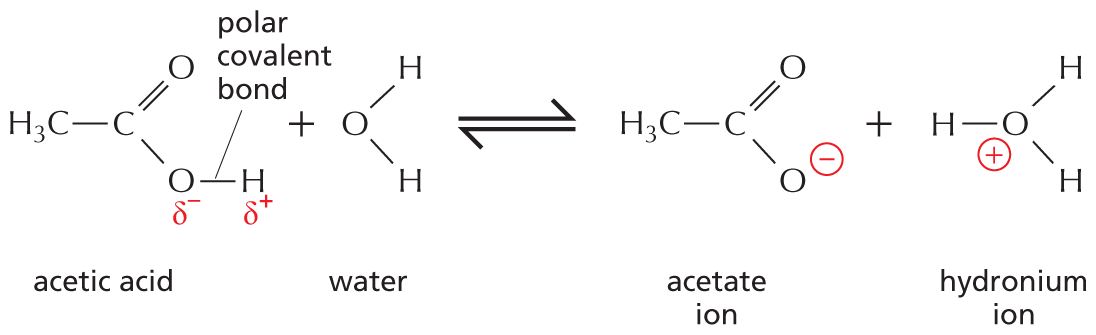
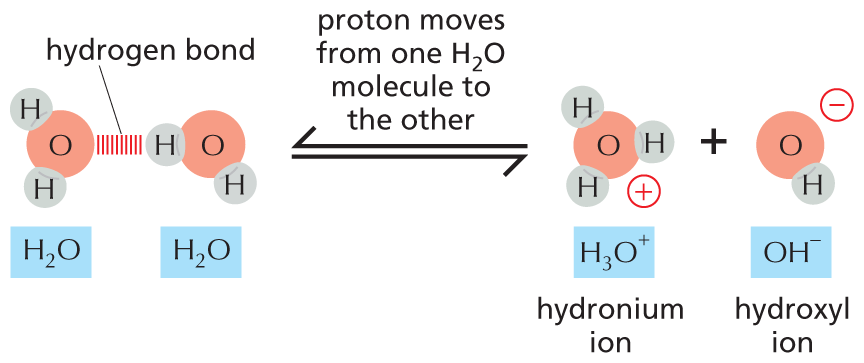
One of the simplest kinds of chemical reaction, and one that has profound significance for cells, takes place when a molecule with a highly polar covalent bond between a hydrogen and another atom dissolves in water. The hydrogen atom in such a bond has given up its electron almost entirely to the companion atom, so it exists as an almost naked positively charged hydrogen nucleus—in other words, a proton (H+). When the polar molecule becomes surrounded by water molecules, the proton will be attracted to the partial negative charge on the oxygen atom of an adjacent water molecule (see Figure 2–11); this proton can thus dissociate from its original partner and associate instead with the oxygen atom of the water molecule, generating a hydronium ion (H3O+) (Figure 2–15A). The reverse reaction—in which a hydronium ion releases a proton—also takes place very readily, so in an aqueous solution, billions of protons are constantly flitting to and fro between one molecule and another.
More information
An equation shows acetic acid reversibly reacting with water to yield an acetate ion and a hydronium ion. The skeletal structures of the reactants and products are shown. Acetic acid contains a central carbon with a single bond to a methyl group, a double bond to an oxygen atom, and a single bond to an oxygen atom with a partial negative charge. The partial negative oxygen atom has a single polar covalent bond to a hydrogen atom with partial positive charge. The water molecule contains an oxygen atom with single bonds to two hydrogen atoms. Acetic acid and water complete the first side of the reaction. On the other side of the reaction, the acetate ion is shown as a central carbon with a single bond to a methyl group, a double bond to an oxygen atom, and a single bond to a negatively charged oxygen ion. The hydronium ion has a central positively charged oxygen ion with a single bond to three hydrogen atoms. Acetic acid reacts with water to form an acetate ion and a hydronium ion.
(A)More information
An equation shows two water molecules reversibly reacting with each other to yield a hydronium ion and a hydroxyl ion. The space filling models on the reactants side shows two water molecules sharing a hydrogen bond between the oxygen atom of the first water molecule and the hydrogen atom of the other water molecule. Both water molecules are labeled with H subscript 2 O. On the other side of the reaction, there is a positively charged hydronium ion shown as an oxygen atom bound to three hydrogen atoms. The hydronium ion is labeled H subscript 3 O superscript plus. The hydroxyl ion has a negative charge and is shown as an oxygen atom bound to a hydrogen atom. The hydroxyl ion is labeled O H superscript minus. Text corresponding to the reverse reaction reads, proton moves from one H subscript 2 O molecule to the other.
(B)Figure 2–15Protons move continuously from one molecule to another in aqueous solutions. (A) The reaction that takes place when a molecule of acetic acid dissolves in water. At pH 7, nearly all of the acetic acid molecules are present as acetate ions. (B) Water molecules are continually exchanging protons with each other to form hydronium and hydroxyl ions. These ions in turn rapidly recombine to form water molecules.
Substances that release protons when they dissolve in water, thus forming H3O+, are termed acids. The higher the concentration of H3O+, the more acidic the solution. Even in pure water, H3O+ is present at a concentration of 10–7 M, as a result of the movement of protons from one water molecule to another (Figure 2–15B). By tradition, the H3O+ concentration is usually referred to as the H+ concentration, even though most protons in an aqueous solution are present as H3O+. To avoid the use of unwieldy numbers, the concentration of H+ is expressed using a logarithmic scale called the pH scale. Pure water has a pH of 7.0 and is thus neutral—that is, neither acidic (pH < 7) nor basic (pH > 7).
QUESTION 2–5
Are there H3O+ ions present in pure water at neutral pH (i.e., at pH = 7.0)? If so, how are they formed?
If they exist, what is the ratio of H3O+ ions to H2O molecules at neutral pH? (Hint: the mass of a water molecule is 18 daltons, and 1 liter of water has a mass of 1 kg.)
Acids are characterized as being strong or weak, depending on how readily they give up their protons to water. Strong acids, such as hydrochloric acid (HCl), lose their protons easily. Acetic acid, on the other hand, is a weak acid because it holds on to its proton fairly tightly when dissolved in water. Many of the acids important in the cell—such as molecules containing a carboxyl (–COOH) group—are weak acids (see Panel 2–2, pp. 72–73). Their tendency to give up a proton with some reluctance is exploited in a wide variety of cellular reactions.
Because protons can be passed readily to many types of molecules in cells, thus altering the molecules’ characters, the H+ concentration inside a cell—its pH—must be closely controlled. Acids will give up their protons more readily if the H+ concentration is low (and the pH is high) and will hold on to their protons (or accept them back) when the H+ concentration is high (and the pH is low).
Molecules that accept protons when dissolved in water are called bases. Just as the defining property of an acid is that it raises the concentration of H3O+ ions by donating a proton to a water molecule, the defining property of a base is that it raises the concentration of hydroxyl (OH–) ions by removing a proton from a water molecule. Sodium hydroxide (NaOH) is basic (or alkaline). NaOH is considered a strong base because it readily dissociates in aqueous solution to form Na+ ions and OH– ions. However, weak bases—which have a weak tendency to accept a proton from water—are more important in cells. Many biologically important weak bases contain an amino (–NH2) group, which can generate OH– by taking a proton from water: –NH2 + H2O ⟶ –NH3+ + OH– (see Panel 2–2, pp. 72–73).
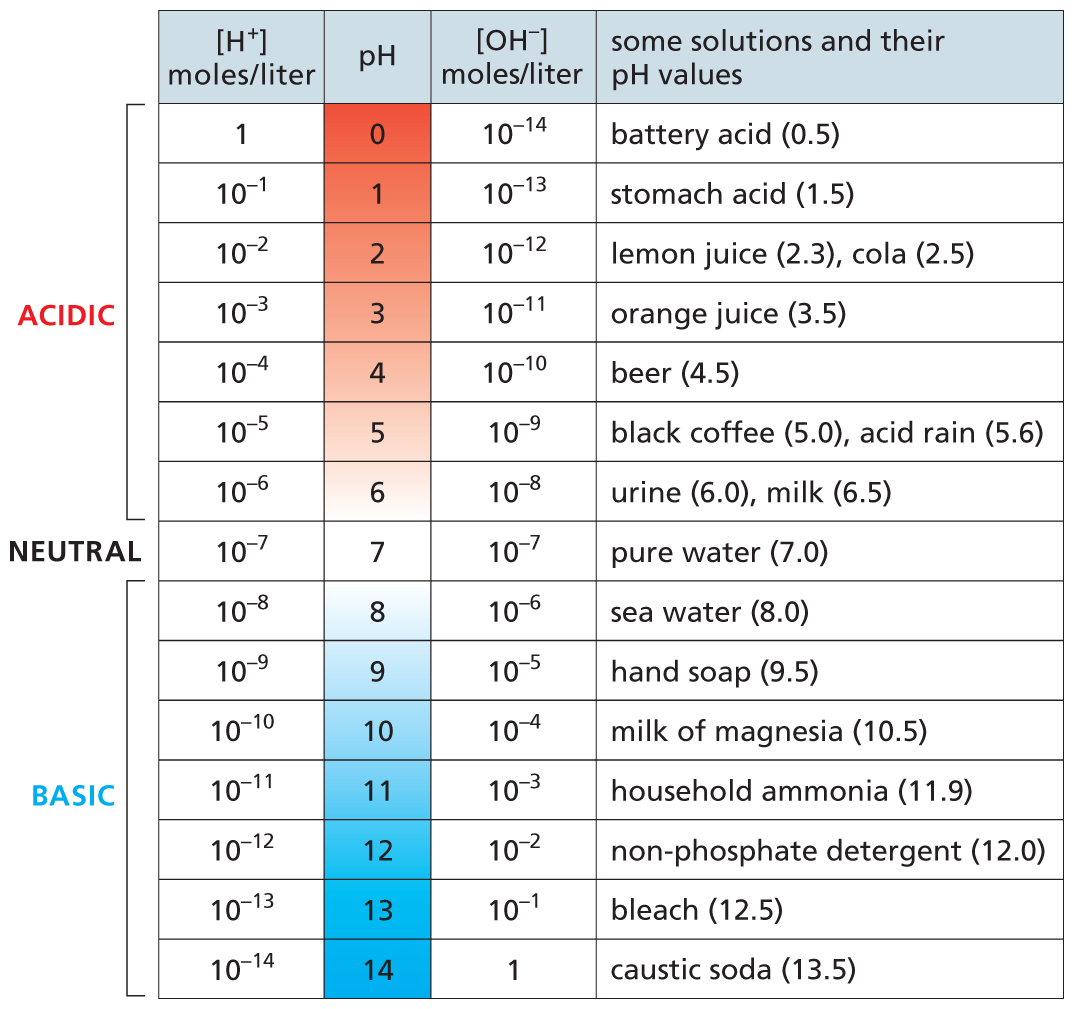
Because an OH– ion combines with a proton to form a water molecule, an increase in the OH– concentration forces a decrease in the H+ concentration, and vice versa (Figure 2–16). A pure solution of water contains an equal concentration (10–7 M) of both ions, rendering it neutral (pH 7). The interior of a cell is kept close to neutral by the presence of buffers. These are mixtures of weak acids and bases that maintain proton concentrations at about pH 7 by releasing protons (acids) or taking them up (bases) whenever the pH changes. This give-and-take keeps the pH of the cell relatively constant under a variety of conditions.
More information
A table shows the p H of common substances arranged in order of increasing p H. The p H values from 0 to 6 are labeled acidic, p H value 7 is labeled neutral, and p H values from 8 to 14 are labeled basic. At each p H value the concentration of hydrogen ions and hydroxyl ions are listed. Examples of solutions near the p H value are also listed.
At p H 0, the concentration of hydronium ions is 1 mole per liter and the concentration of hydroxyl ions is 10 to the negative 14th power moles per liter. Battery acid has a p H of 0.5. At p H 1, the concentration of hydronium ions is 10 to the negative 1st power moles per liter and the concentration of hydroxyl ions is 10 to the negative 13th power moles per liter. Stomach acid has a p H of 1.5. At p H 2, the concentration of hydronium ions is 10 to the negative 2nd power moles per liter and the concentration of hydroxyl ions is 10 to the negative 12th power moles per liter. Lemon juice has a p H of 2.3 and cola has a p H of 2.5 At p H 3, the concentration of hydronium ions is 10 to the negative 3rd power moles per liter and the concentration of hydroxyl ions is 10 to the negative 11th power moles per liter. Orange juice has a p H of 3.5. At p H 4, the concentration of hydronium ions is 10 to the negative 4th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 10th power moles per liter. Beer has a p H of 4.5. At p H 5, the concentration of hydronium ions is 10 to the negative 5th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 9th power moles per liter. Black coffee has a p H of 5 and acid rain has a p H of 5.6. At p H 6, the concentration of hydronium ions is 10 to the negative 6th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 8th power moles per liter. Urine has a p H of 6 and milk has a p H of 6.5. At p H 7, the concentration of hydronium ions is 10 to the negative 7th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 7th power moles per liter. Pure water has a p H of 7. At p H 8, the concentration of hydronium ions is 10 to the negative 8th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 6th power moles per liter. Sea water has a p H of 8. At p H 9, the concentration of hydronium ions is 10 to the negative 9th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 5th power moles per liter. Hand soap has a p H of 9.5. At p H 10, the concentration of hydronium ions is 10 to the negative 10th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 4th power moles per liter. Milk of magnesia has a p H of 10.5. At p H 11, the concentration of hydronium ions is 10 to the negative 11th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 3rd power moles per liter. Household ammonia has a p H of 11.9. At p H 12, the concentration of hydronium ions is 10 to the negative 12th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 2nd power moles per liter. Non-phosphate detergent has a p H of 12. At p H 13, the concentration of hydronium ions is 10 to the negative 13th power moles per liter and the concentration of hydroxyl ions is 10 to the negative 1st power moles per liter. Bleach has a p H of 12.5. At p H 14, the concentration of hydronium ions is 10 to the negative 14th power moles per liter and the concentration of hydroxyl ions is 1 mole per liter. Caustic soda has a p H of 13.5.
Figure 2–16In aqueous solutions, the concentration of hydroxyl (OH–) ions increases as the concentration of H3O+ (or H+) ions decreases. The product of the two values, [OH–] × [H+], is always 10–14 (moles/liter)2. At neutral pH, [OH–] = [H+], and both ions are present at 10–7 M. Also shown are examples of common solutions along with their approximate pH values.
The smallest particle of an element that still retains its distinctive chemical properties; consists of a positively charged nucleus surrounded by a cloud of negatively charged electrons.
A weak noncovalent interaction between a positively charged hydrogen atom in one molecule and a negatively charged atom, such as nitrogen or oxygen, in another; hydrogen bonds are key to the structure and properties of water.
Nonpolar, uncharged molecule or part of a molecule that forms no hydrogen bonds with water molecules and therefore does not dissolve; literally, “water fearing.”
Chemical association that does not involve the sharing of electrons; singly they are relatively weak, but they can sum together to produce strong, highly specific interactions between molecules. Examples are hydrogen bonds and van der Waals attractions.
Force that draws together oppositely charged atoms. Examples include ionic bonds and the attractions between molecules containing polar covalent bonds.
A noncovalent interaction that forces together the hydrophobic portions of dissolved molecules to minimize their disruption of the hydrogen-bonded network of water; causes membrane phospholipids to self-assemble into a bilayer and helps to fold proteins into a compact, globular shape.