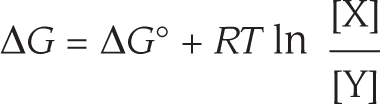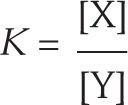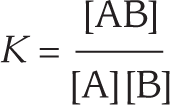Life depends on the highly specific chemical reactions that take place inside cells. The vast majority of these reactions are catalyzed by proteins called enzymes. Enzymes, like cells, must obey the second law of thermodynamics. Although an individual enzyme can greatly accelerate an energetically favorable reaction—one that produces disorder in the universe—it cannot force an energetically unfavorable reaction to occur. Cells, however, must do just that in order to grow and divide—or just to survive. They must build highly ordered and energy-rich molecules from small and simple ones—a process that requires an input of energy.
To understand how enzymes promote the acceleration of the specific chemical reactions needed to sustain life, we first need to examine the energetics involved. In this section, we consider how the free energy of molecules contributes to their chemistry, and we see how changes in free energy—which reflect how much total disorder is generated in the universe by a reaction—influence whether and how a reaction will proceed. Examining these energetic concepts will reveal how enzymes working together can harness the free-energy changes of chemical reactions to drive the energetically unfavorable reactions that produce biological order. This type of enzyme-assisted catalysis is crucial for cells: without it, life could not exist.
Chemical Reactions Proceed in the Direction That Causes a Loss of Free Energy
Paper burns readily, releasing into the atmosphere water and carbon dioxide as gases, while simultaneously releasing energy as heat:
paper + O2⟶ smoke + ashes + heat + CO2 + H2O
QUESTION 3–2
In which of the following reactions does the red atom undergo an oxidation?
Na⟶ Na+ (Na atom ⟶ Na+ ion)
Cl⟶ Cl– (Cl atom ⟶ Cl– ion)
CH3CH2OH ⟶ CH3CHO (ethanol ⟶ acetaldehyde)
CH3CHO ⟶ CH3COO– (acetaldehyde ⟶ acetic acid)
CH2=CH2⟶ CH3CH3 (ethene ⟶ ethane)
This reaction occurs in only one direction: smoke and ashes never spontaneously gather carbon dioxide and water from the heated atmosphere and reconstitute themselves into paper. When paper burns, most of its chemical energy is dissipated as heat. This heat is not lost from the universe, since energy can never be created or destroyed; instead, it is irretrievably dispersed in the chaotic random thermal motions of molecules. In the language of thermodynamics, there has been a release of free energy—that is, energy that can be harnessed to do work or drive chemical reactions. This release reflects a change in the way the energy and molecules had been stored in the paper; the greater the free-energy change, the greater the amount of disorder created in the universe when the reaction occurs.
We will discuss free energy in more detail shortly, but a general principle can be summarized as follows: chemical reactions proceed only in the direction that leads to a loss of free energy. In other words, the spontaneous direction for any reaction is the direction that goes “downhill.” A “downhill” reaction in this sense is said to be energetically favorable.
Enzymes Reduce the Energy Needed to Initiate Spontaneous Reactions
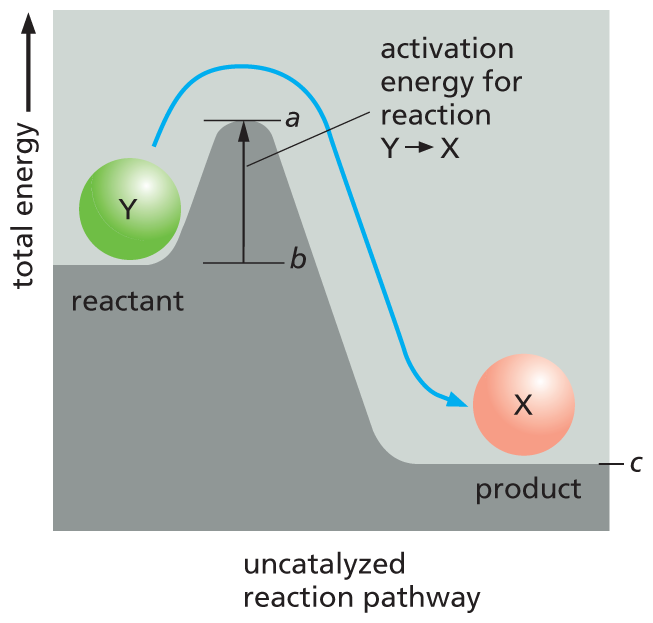
Although the most energetically favorable form of carbon under ordinary conditions is CO2, and that of hydrogen is H2O, a living organism will not disappear in a puff of smoke, and the book in your hands will not burst spontaneously into flames, as suggested by the previous incendiary reaction. This is because the molecules in both the living organism and the book are in a relatively stable state, and they cannot be changed to lower-energy states without an initial input of energy. In other words, a molecule requires a boost over an energy barrier before it can undergo a chemical reaction that moves it to a lower-energy (more stable) state. This boost is known as the activation energy (Figure 3–12A). In the case of a burning book, the activation energy is provided by the heat of a lighted match.
More information
A graph depicts the uncatalyzed reaction pathway. Total energy is plotted along the vertical axis. The curve shows that reactant Y is at an energy level b initially; b corresponds to about half of the total energy, after which it ascends to a peak energy level a; ‘a’ corresponds to about three-fourths of the total energy. After ‘a’ the total energy level descends to a minimum value c which corresponds to energy level of product X. c corresponds to about one-fourth of the total energy. An upward arrow marks the vertical distance between b to a as activation energy for reaction converting Y to X.
(A)More information
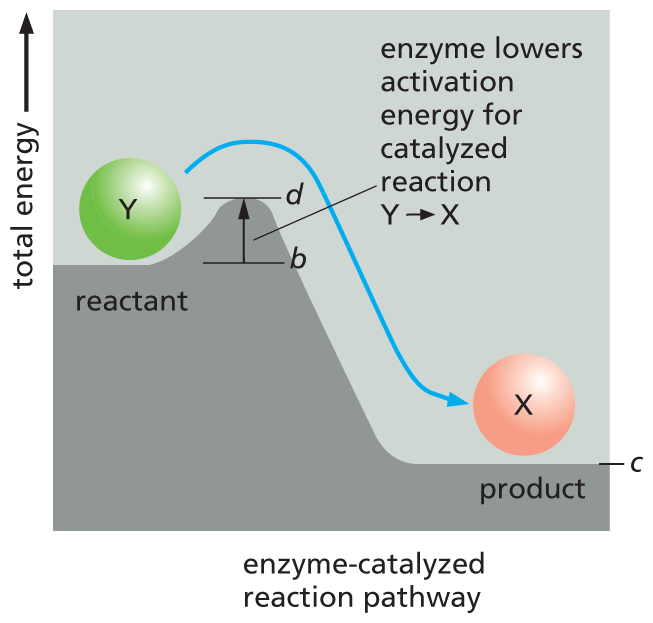
A graph depicts the reaction pathways of enzyme-catalyzed. Enzyme-catalyzed reaction pathway: The curve shows that the reactant Y is at energy level b initially, after which it ascends to a smaller peak value d (d is less than a); after d, the total energy descends to a minimum value c. An upward arrow marks the vertical distance between b and d with a corresponding text that reads, enzyme lowers activation energy for catalyzed reaction, converting Y to X.
(B)Figure 3–12Even energetically favorable reactions require activation energy to get them started. (A) Compound Y (a reactant) is in a relatively stable state; thus energy is required to convert it to compound X (a product), even though X is at a lower overall energy level than Y. This conversion will not take place, therefore, unless compound Y can acquire enough activation energy (energy a minus energy b) from its surroundings to undergo the reaction that converts it into compound X. This energy may be provided by means of an unusually energetic collision with other molecules. For the reverse reaction, X ⟶ Y, the activation energy required will be much larger (energy a minus energy c); this reaction will therefore occur much more rarely. The total energy change for the energetically favorable reaction Y ⟶ X is energy c minus energy b, a negative number, which corresponds to a loss of free energy. (B) Energy barriers for specific reactions can be lowered by catalysts, as indicated by the line marked d. Enzymes are particularly effective catalysts because they greatly reduce the activation energy for the reactions they catalyze. Note that activation energies are always positive and that the enzyme does not change the ΔG (the free-energy difference between the product and reactant) of the reaction.
QUESTION 3–3
The enzyme carbonic anhydrase is one of the speediest enzymes known. It catalyzes the rapid conversion of CO2 gas into the much more soluble bicarbonate ion (HCO3–). The reaction:
CO2 + H2O ↔ HCO3– + H+
is very important for the efficient transport of CO2 from tissues, where CO2 is produced by respiration, to the lungs, where it is exhaled. Carbonic anhydrase accelerates this reaction 107-fold, hydrating 105 CO2 molecules per second at its maximal speed. A more typical enzyme produces around 1000 product molecules per second. Compare carbonic anhydrase with this second, more mainstream enzyme using two diagrams like the one shown in Figure 3–13. Explain how your two diagrams have been designed to explain the 100-fold difference between the two maximum rates.
Cells don’t raise their temperature to drive biological reactions; instead, that assistance is provided by enzymes. Each enzyme binds tightly to one or two molecules, called substrates, and holds them in a way that greatly reduces the activation energy needed to facilitate a specific chemical reaction (Figure 3–12B). A substance that can lower the activation energy of a reaction is termed a catalyst; catalysts boost the rate of chemical reactions because they allow a much larger proportion of the random collisions with surrounding molecules to kick the substrates over the energy barrier, as illustrated in Figure 3–13 and Figure 3–14A. Enzymes are among the most effective catalysts known. They can speed up reactions by a factor of as much as 1014—that is, trillions of times faster than the same reactions would proceed without an enzyme catalyst. Enzymes therefore allow reactions that would not otherwise occur to proceed rapidly at the normal temperature inside cells.
More information
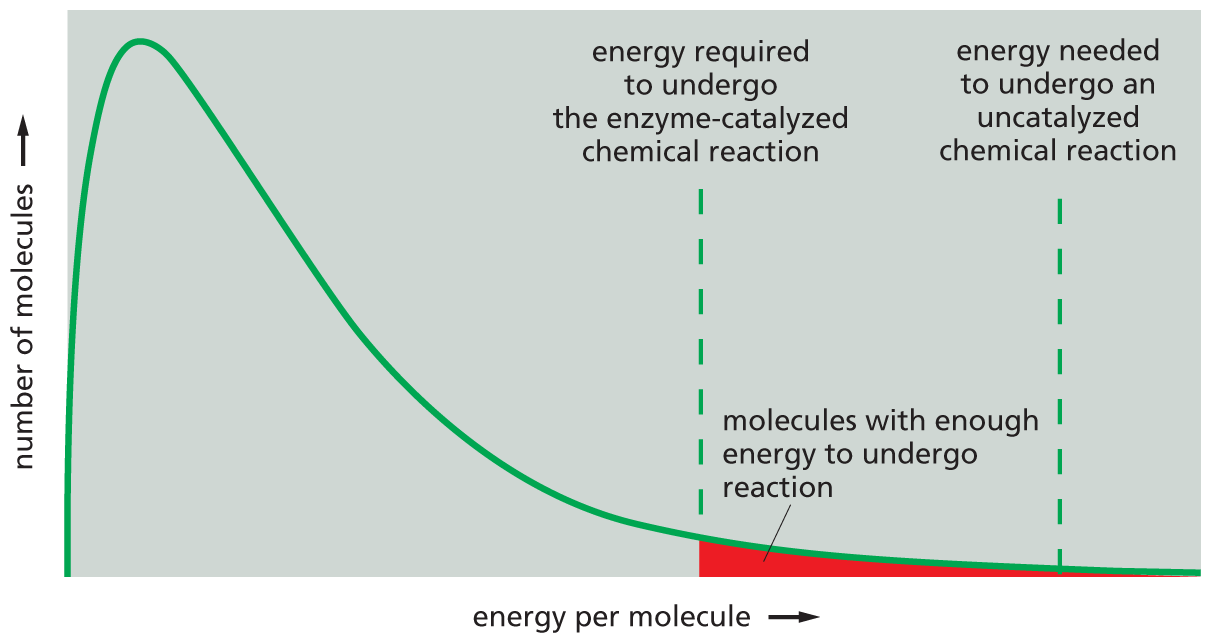
A graph shows that lowering the activation energy increases the probability of a reaction occurring. The horizontal axis plots energy per molecule and the vertical axis plots number of molecules. The graph shows a curve which starts from the origin and increases steeply to a peak value after which it descends gradually. A vertical dashed line originates from a point which is a little greater than half of the horizontal axis; this line represents the energy required to undergo the enzyme-catalyzed chemical reaction. Another vertical dashed line originates from a point which is a little greater than three-fourths of the horizontal axis and represents the energy needed to undergo an unanalyzed chemical reaction. The area under the curve between these two dashed lines is shaded and represents molecules with enough energy to undergo a reaction.
Figure 3–13Lowering the activation energy greatly increases the probability that a reaction will occur. At any given instant, a population of identical substrate molecules will have a range of energies, distributed as shown on the graph. The varying energies come from collisions with surrounding molecules, which make the substrate molecules jiggle, vibrate, and spin. Because the vast majority of molecules have nowhere near the energy required to undergo a chemical reaction (dashed lines), most biological reactions will almost never happen without enzyme catalysis. Even when an enzyme lowers the activation energy of the reaction, only a small fraction of substrate molecules will experience the highly energetic collisions (red-shaded area) needed to reach an energy state high enough for them to undergo a reaction.
More information
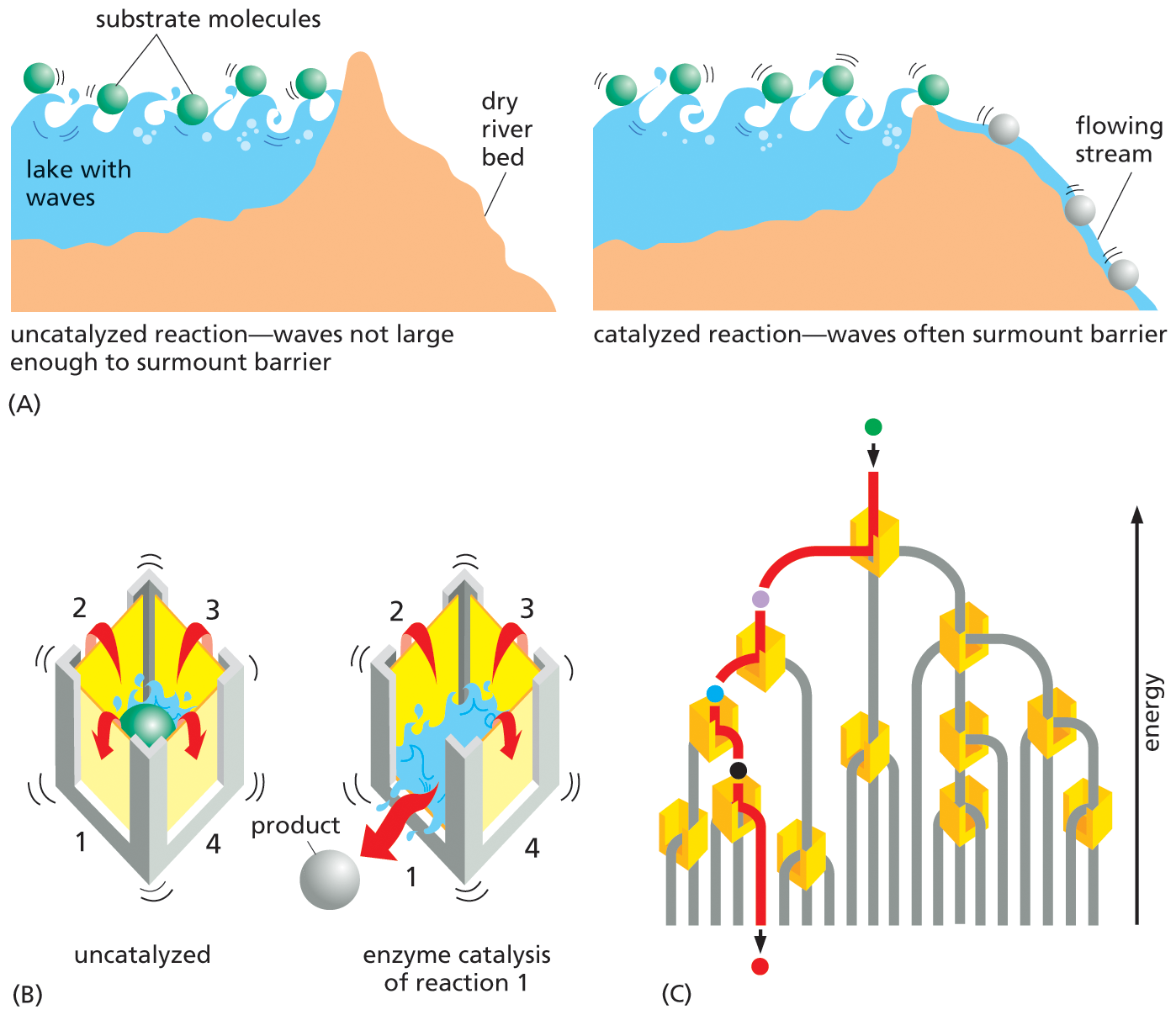
A three-part illustration depicts lowering of the activation energy barrier in reactions catalyzed by enzymes, and the energy changes in sequential enzymatic reaction pathways. Part A: An illustration uses water to demonstrate the lowering of activation energy barrier in reactions catalyzed by enzymes. Part B: An illustration uses a box to depict the lowering of activation energy barrier in reactions catalyzed by enzymes. Part C: An illustration shows the energy changes in sequential enzymatic reaction pathways.
Figure 3–14Enzymes catalyze reactions by lowering the activation-energy barrier. (A) The dam represents the activation energy, which is lowered by enzyme catalysis. Each green ball represents a potential substrate molecule that is bouncing up and down in energy level owing to constant encounters with waves, an analogy for the thermal bombardment of substrate molecules by surrounding water molecules. When the barrier—the activation energy—is lowered significantly, the balls (substrate molecules) with sufficient energy can roll downhill, an energetically favorable movement. (B) The four walls of the box represent the activation-energy barriers for four different chemical reactions that are all energetically favorable because the products are at lower energy levels than the substrates. In the left-hand box, none of these reactions occurs because even the largest waves are not large enough to surmount any of the energy barriers. In the right-hand box, enzyme catalysis lowers the activation energy for reaction number 1 only; now the jostling of the waves allows the substrate molecule to pass over this energy barrier, allowing reaction 1 to proceed (Movie 3.1). (C) A branching set of reactions with a selected set of enzymes (yellow boxes) serves to illustrate how a series of enzyme-catalyzed reactions—by controlling which reaction will take place at each junction—determines the exact reaction pathway followed by each molecule inside the cell.
Unlike the effects of temperature, enzymes are highly selective. Each enzyme usually speeds up—or catalyzes—only one particular reaction out of the several possible reactions that its substrate molecules could undergo. In this way, enzymes direct each of the many different molecules in a cell along specific reaction pathways (Figure 3–14B and C), thereby producing the compounds that the cell actually needs.
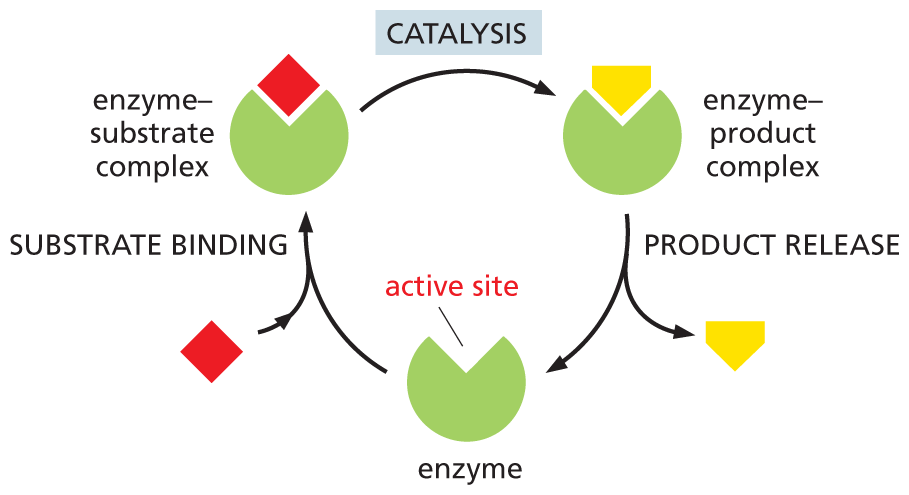
Like all catalysts, enzyme molecules themselves remain unchanged after participating in a reaction and can therefore act over and over again (Figure 3–15). In Chapter 4, we will discuss further how enzymes work, after we have looked in detail at the molecular structure of proteins.
More information
An illustration shows the cycle of catalysis. An enzyme binds with a substrate molecule into the substrate binding site of the enzyme to form an enzyme substrate complex. The enzyme-substrate complex undergoes catalysis to form an enzyme-product complex following which the product molecule is released. The active site of the enzyme is empty now, ready to bind with the substrate again and thus the cycle continues.
Figure 3–15Enzymes convert substrates to products. Catalysis takes place in a cycle in which a substrate molecule (red) binds to an enzyme and undergoes a reaction to form a product molecule (yellow), which then gets released. Although the enzyme participates in the reaction, it emerges from the process in the same state in which it began; in other words, the enzyme itself is not permanently altered by the reaction. It can therefore function over and over again. (Dynamic Figure) The enzyme can therefore perform the same reaction, with new substrate molecules, over and over again (below).
The Free-Energy Change for a Reaction Determines Whether It Can Occur Spontaneously
According to the second law of thermodynamics, a chemical reaction can proceed only if it results in a net (overall) increase in the disorder of the universe. As discussed earlier, this disorder increases when “useful energy” that could be harnessed to do work is dissipated as heat (see Figure 3–5). The useful energy in a system is known as its free energy, or G. But because chemical reactions involve a transition from one molecular state to another, the quantity that most interests chemists and cell biologists is actually the free-energy change, denoted as ΔG (“Delta G”).
For any collection of molecules, ΔG denotes the amount of disorder created in the universe when a reaction involving these molecules takes place. Energetically favorable reactions, by definition, are those that create disorder in the universe by decreasing the free energy of the system to which they belong; in other words, they have a negativeΔG (Figure 3–16).
More information
An illustration shows the free energy change in energetically favorable and unfavorable reactions. In an energetically favorable reaction, a molecule Y is converted into molecule X. Text reads, this reaction can occur spontaneously; the free energy of Y is greater than the free energy of X. Therefore delta G is negative (less than 0), and the disorder of the universe increases when Y is converted to X.
In an energetically unfavorable reaction, molecule X is converted back to molecule Y. Text reads, this reaction can occur only if it is driven by being coupled to a second, energetically favorable reaction; if the reaction X to Y occurred, delta G would be positive (greater than 0), and the universe would become more ordered.
Figure 3–16Energetically favorable reactions have a negative ΔG, whereas energetically unfavorable reactions have a positive ΔG. Imagine, for example, that molecule Y has a free energy (G) of 10 kilojoules (kJ) per mole, whereas X has a free energy of 4 kJ/mole. The reaction Y ⟶ X therefore has a ΔG of –6 kJ/mole, making it energetically favorable.
A reaction can occur spontaneously only if ΔG is negative. On a macroscopic scale, an example of an energetically favorable reaction with a negative ΔG is the relaxation of a compressed spring into an expanded state, which releases its stored elastic energy as heat to its surroundings. On a microscopic scale, an energetically favorable reaction—one with a negative ΔG—occurs when salt (NaCl) dissolves in water. Note that just because a reaction can occur spontaneously does not mean it will occur quickly. Although the oxidation of glucose to CO2 and H2O is energetically highly favorable, it would take centuries to complete without the enzymes cells use to speed up the process. The decay of diamonds into graphite is also a spontaneous process—but it takes millions of years!
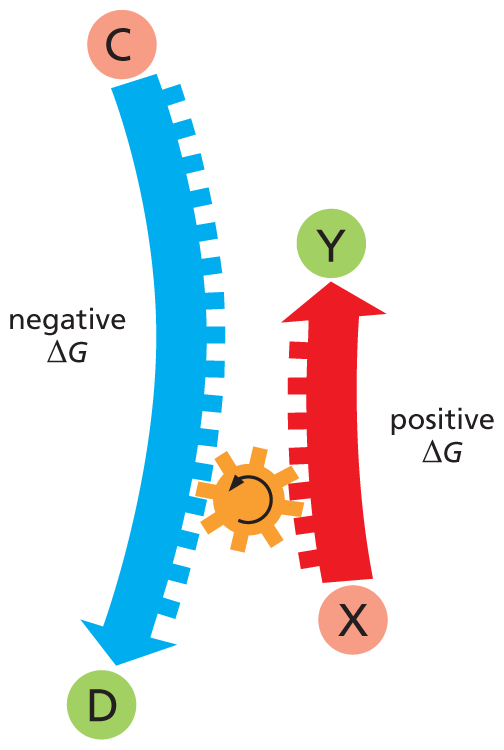
Energetically unfavorable reactions, by contrast, create order in the universe; they have a positiveΔG. Such reactions—for example, the formation of a peptide bond between two amino acids—cannot occur spontaneously; they take place only when they are coupled to a second reaction with a negative ΔG large enough that the net ΔG of the entire process is negative (Figure 3–17). Life is possible because enzymes can create biological order by coupling energetically unfavorable reactions with energetically favorable ones. These critical concepts are summarized, with examples, in Panel 3–1 (pp. 100–101).
More information
An illustration shows reaction coupling in an energetically unfavorable reaction. A reaction sequence represented by a downward arrow converts molecule C to product D; this reaction has a negative delta G. Another reaction represented by an upward arrow converts molecule X to product Y; this reaction has a positive delta G. These two reactions are coupled with a gear cog such that as one arrow moves down the other arrow moves up.
Figure 3–17Reaction coupling can drive an energetically unfavorable reaction. The energetically unfavorable (ΔG > 0) reaction X ⟶ Y cannot occur unless it is coupled to an energetically favorable (ΔG < 0) reaction C ⟶ D, such that the net free-energy change for the pair of reactions is negative (less than 0).
ΔG Changes as a Reaction Proceeds Toward Equilibrium
It’s easy to see how a tensed spring, when left to itself, will relax and release its stored energy to the environment as heat. But chemical reactions are a bit more complex—and harder to intuit. That’s because whether a reaction will proceed in a particular direction depends not only on the energy stored in each individual molecule, but also on the concentrations of the molecules in the reaction mixture. Going back to our jiggling box of coins, more coins will flip from a head to a tail orientation when the box contains 90 heads and 10 tails than when the box contains 10 heads and 90 tails.
The same is true for a chemical reaction. As the energetically favorable reaction Y ⟶ X proceeds, the concentration of the product X will increase and the concentration of the substrate Y will decrease. This change in relative concentrations of substrate and product will cause the ratio of Y to X to shrink, making the initially favorable ΔG less and less negative. Unless more Y is added, the reaction will slow and eventually stop.
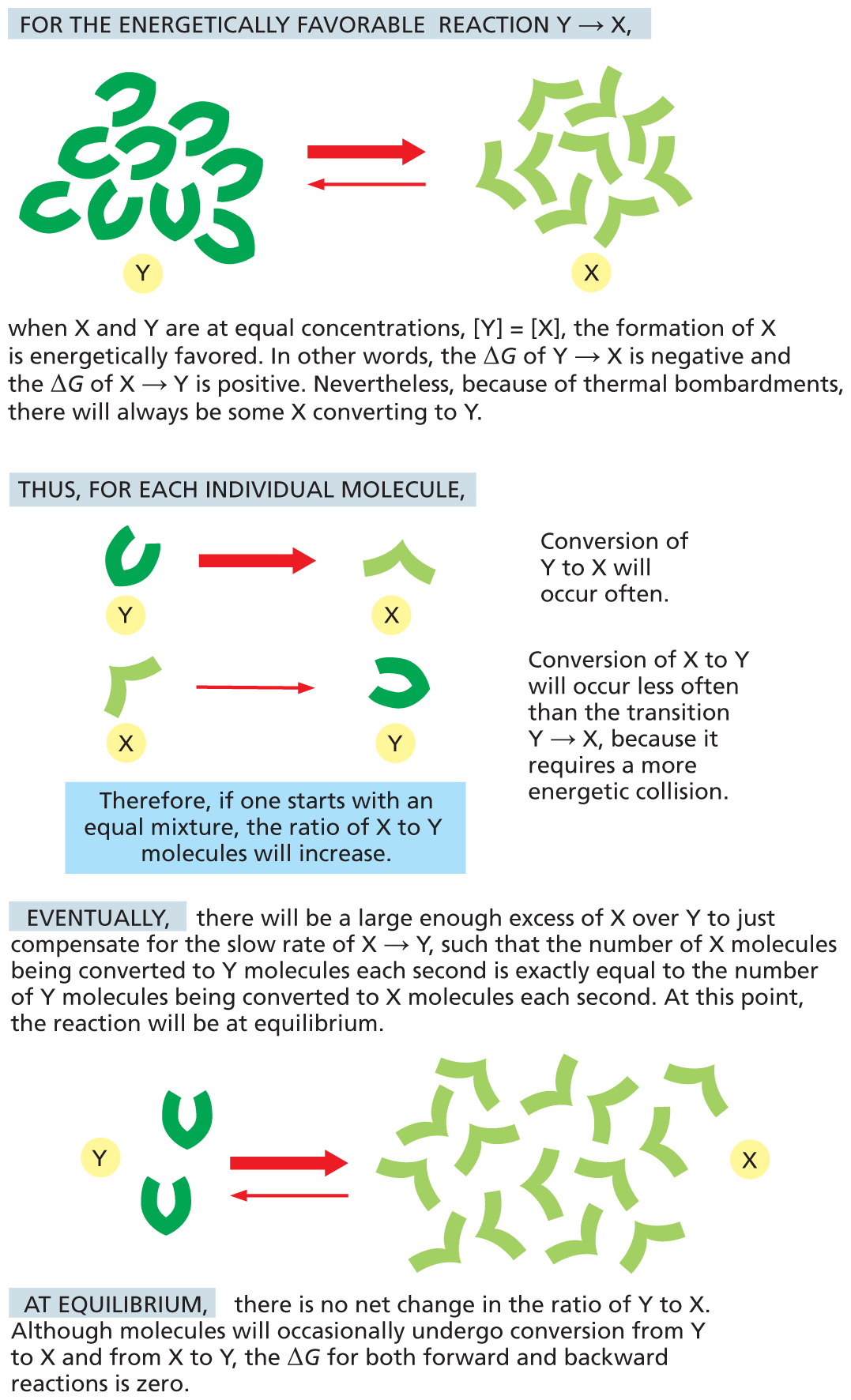
Because ΔG changes as products accumulate and substrates are depleted, chemical reactions will generally proceed until they reach a state of equilibrium. At that point, the rates of the forward and reverse reactions are equal, and there is no further net change in the concentrations of substrate or product (Figure 3–18). For reactions at chemical equilibrium, ΔG = 0, so the reaction will not proceed forward or backward, and no work can be done.
More information
An illustration shows how reactions reach chemical equilibrium. For an energetically favorable reaction involving the conversion of molecule Y to X, a cluster of Y molecules are converted into a cluster of X molecules. Text reads, when X and Y are at equal concentrations (concentration of Y equals concentration of X), the formation of X is energetically favored. In other words, the delta G of Y to X is negative and the delta G of X to Y is positive. Nevertheless because of thermal bombardments, there will always be some X converting to Y.
Thus for each individual molecule, conversion of Y to X will occur often. A schematic shows a molecule of Y being converted to a molecule of X. Conversion of X to Y will occur less often than the transition Y to X because it requires a more energetic collision. A schematic shows a molecule of X being converted to a molecule of Y. Text reads, therefore, if one starts with an equal mixture, the ratio of X and Y molecules will increase. Eventually, there will be a large enough excess of X over Y to just compensate for the slow rate of X to Y, such that the number of X molecules being converted to Y molecules each second is exactly equal to the number of Y molecules being converted to X molecules each second. At this point, the reaction will be at equilibrium.
A schematic illustration below shows two molecules of Y being converted to many molecules of X; this reaction can be reversed but at a smaller rate. Text reads, At equilibrium, there is no net change in the ratio of Y to X. Although molecules will occasionally undergo conversion from Y to X and X to Y, the delta G for both forward and backward reactions is zero.
Figure 3–18Reactions will eventually reach a chemical equilibrium. At that point, the forward and the backward fluxes of reacting molecules are equal and opposite. The width of each arrow indicates the relative probability that an individual molecule will undergo the indicated chemical reaction.
Such a state of chemical inactivity would be incompatible with life, inevitably allowing chemical decay to overcome the cell. Living cells therefore expend energy to avoid reaching a state of complete chemical equilibrium. They are constantly exchanging materials with their environment: replenishing nutrients and eliminating waste products. In addition, many of the individual reactions in the cell’s complex metabolic network also exist in disequilibrium because the products of one reaction are continually being siphoned off to become the substrates in a subsequent reaction. Rarely do products and substrates reach concentrations at which the forward and reverse reaction rates are equal.
The Standard Free-Energy Change, ΔG°, Makes It Possible to Compare the Energetics of Different Reactions
Because ΔG depends on the concentrations of the molecules in the reaction mixture at any given time, it is not a particularly useful value for comparing the relative energetics of different types of chemical reactions. But such assessments are necessary, for example, to predict whether an energetically favorable reaction is likely to have a ΔG negative enough to drive an energetically unfavorable reaction. To compare reactions in this way, we need to turn to the standard free-energy change, ΔG°, of the reaction. A reaction’s ΔG° is independent of concentration; it depends only on the intrinsic characters of the reacting molecules, based on their behavior under ideal conditions where the concentrations of all the reactants are set to the same fixed value of 1 mole/liter in aqueous solution. Although these ideal conditions are rarely present in cells, comparing the ΔG° of different metabolic reactions provides a sense of their intrinsic favorability (see Panel 3–1, p. 100).
The ΔG of a reaction can be calculated from ΔG° if the concentrations of the reactants and products are known. For the simple reaction Y ⟶ X, their relationship follows this equation:
More information
An equation shows that delta G equals delta G naught plus R T times the natural log of the concentration of x divided by the concentration of y.
where ΔG° is in kilojoules per mole, [Y] and [X] denote the concentrations of Y and X in moles/liter (a mole is 6 × 1023 molecules of a substance), ln is the natural logarithm, and RT is the product of the gas constant, R, and the absolute temperature, T. (At 37°C, RT = 2.58 kJ/mole.)
QUESTION 3–4
Consider the analogy of the jiggling box containing coins that was described on p. 87. The reaction, the flipping of coins that either face heads up (H) or tails up (T), is described by the equation H ↔ T, where the rate of the forward reaction equals the rate of the reverse reaction.
What are ΔG and ΔG° in the analogy?
What corresponds to the temperature at which the reaction proceeds? What corresponds to the activation energy of the reaction? Assume you have an “enzyme,” called jigglase, which catalyzes this reaction. What would the effect of jigglase be and what, mechanically, might jigglase do in this analogy?
From this equation, we can see that when the concentrations of reactants and products are equal—in other words, [X]/[Y] = 1—the value of ΔG equals the value of ΔG° (because ln 1 = 0). Thus when the reactants and products are present in equal concentrations, the direction of the reaction depends entirely on the intrinsic properties of the molecules.
The Equilibrium Constant Is Directly Proportional to ΔG°
Although chemical reactions rarely reach equilibrium inside the cell, they do proceed in that direction. Thus for any given reaction, knowing where the equilibrium point lies will reveal which way that reaction will proceed—and how far it will go. For example, if a reaction is at equilibrium when the concentration of the product is 10 times the concentration of the substrate, and a cell contains a surplus of substrate and little or no product, the reaction will proceed forward, and it may eventually reach equilibrium. The ratio of product to substrate at this equilibrium point is called the reaction’s equilibrium constant, K.
For the simple reaction Y ⟶ X,
More information
An equation shows that K equals the concentration of x divided by the concentration of y.
where [X] is the concentration of the product and [Y] is the concentration of the substrate at equilibrium. In the example we just described, K = 10.
The equilibrium constant depends on the intrinsic properties of the molecules involved, just like ΔG° does. In fact, the equilibrium constant is directly proportional to ΔG°.
ΔG° = –5.94 log K
Thus, for the reaction Y ⟶ X, where K = 10, ΔG° = –5.94 kJ/mole. In fact, for every 5.94-kJ/mole difference in free energy at 37°C, the equilibrium constant for a reaction changes by a factor of 10, as shown in Table 3–1. Thus, the more energetically favorable the reaction, the more product will accumulate as the reaction proceeds to equilibrium. For a reaction with a ΔG° of –17.8 kJ/mole, K will equal 1000, which means that at equilibrium, there will be 1000 molecules of product for every molecule of substrate present. To learn more about the relationship between K and ΔG°, see Panel 3–1 (p. 100).
TABLE 3–1 RELATIONSHIP BETWEEN THE STANDARD FREE-ENERGY CHANGE, ∆G°, AND THE EQUILIBRIUM CONSTANT
Equilibrium Constant, K ([X]/[Y])
Standard Free-Energy Change (∆G°) for Reaction Y ⟶ X (kJ/mole)
105
–29.7
104
–23.8
103
–17.8
102
–11.9
101 (= 10)
–5.9
100 (= 1)
0
10–1
5.9
10–2
11.9
10–3
17.8
10–4
23.8
10–5
29.7
Values of the equilibrium constant were calculated for the simple chemical reaction Y ⟶ X, using the equation given in the text.
The ∆G° values given here are in kilojoules per mole at 37°C. As explained in the text, ∆G° represents the free-energy difference under standard conditions (where all components are present at a concentration of 1 mole/liter).
From this table, we see that if there is a favorable free-energy change of –17.8 kJ/mole for the transition Y ⟶ X, there will be 1000 times more molecules of X than of Y at equilibrium (K = 1000).
In Complex Reactions, the Equilibrium Constant Includes the Concentrations of All Reactants and Products
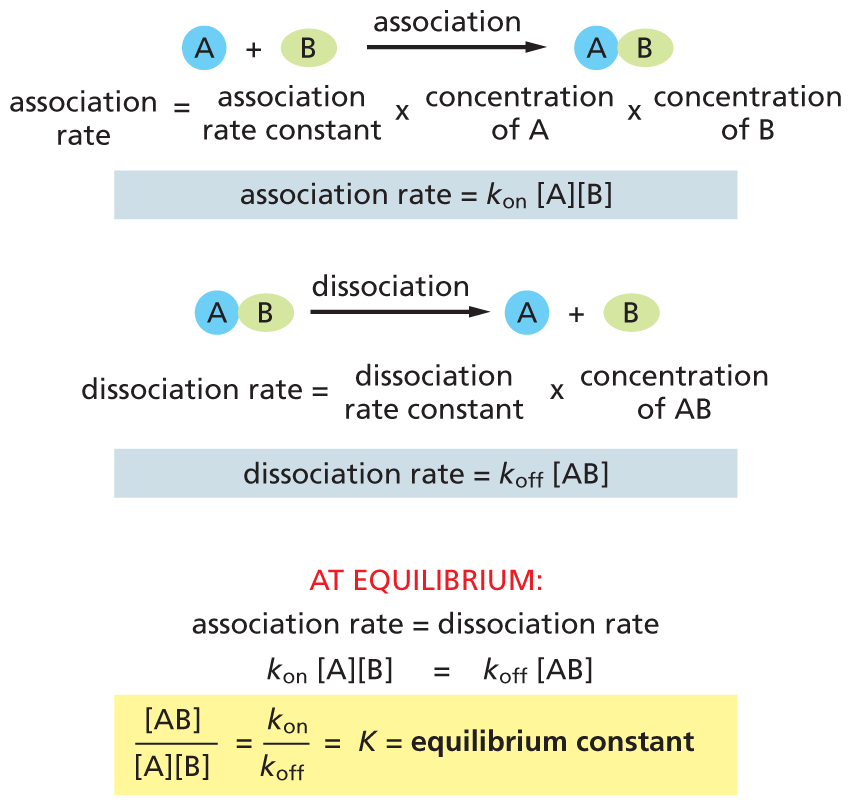
We have so far discussed the simplest of reactions, Y ⟶ X, in which a single substrate is converted into a single product. But inside cells, it is more common for two reactants to combine to form a single product: A + B ⟶ AB. How can we predict how this reaction will proceed?
The same principles apply, except that in this case the equilibrium constant K includes the concentrations of both of the reactants, in addition to the concentration of the product:
More information
An equation shows that K equals the concentration of A B divided by the multiplication of concentration of A and concentration B.
The concentrations of both reactants are multiplied in the denominator because the formation of product AB depends on the collision of A and B, and these encounters occur at a rate that is proportional to [A] × [B] (Figure 3–19). As with single-substrate reactions, ΔG° = –5.94 log K at 37°C. Thus, the relationship between K and ΔG° is the same as that shown in Table 3–1.
More information
An equation shows that delta G naught equals negative 5.94 times the log of K. Reactions are given as follows:
A plus B undergoes association to form A B as product.
Association rate equals association rate constant multiplied by concentration A multiplied by concentration B.
Association rate equals k subscript on concentration of A times concentration of B.
A B undergoes dissociation to form yields A plus B as products.
Dissociation rate equals dissociation rate constant multiplied by concentration of A B.
Dissociation rate equals k subscript off times the concentration of A B.
At equilibrium:
Association rate equals Dissociation rate k subscript on times the concentration of A times concentration of B which equals k subscript off times concentration of A B
Concentration of A B over concentration of A times concentration of B equals K which equals equilibrium constant.
Figure 3–19The equilibrium constant, K, for the reaction A + B ⟶ AB depends on the concentrations of A, B, and AB. Molecules A and B must collide in order to interact, and the association rate is therefore proportional to the product of their individual concentrations [A] × [B]. As shown, the ratio of the rate constants kon and koff for the association (bond formation) and the dissociation (bond breakage) reactions, respectively, is equal to the equilibrium constant, K.
PANEL 3–1FREE ENERGY AND CATALYSIS
FREE ENERGY
This panel reviews the concept of free energy and offers examples showing how changes in free energy determine whether—and how—biological reactions occur.
The molecules of a living cell possess energy because of their vibrations, rotations, and movement through space, and because of the energy that is stored in the bonds between individual atoms.
More information
A 9 panel illustration shows examples of free energy and catalysis reactions. These panels contain text and illustrations. The titles of the panels are free energy, reactions cause disorder, predicting reactions, delta G, spontaneous reactions, reaction rates, delta G equals zero at chemical equilibrium, coupled reactions, and high-energy bonds.
The free energy, G (in kJ/mole), measures the energy of a molecule that could in principle be used to do useful work at constant temperature, as in a living cell. Energy can also be expressed in calories (1 joule = 0.24 calories).
REACTIONS CAUSE DISORDER
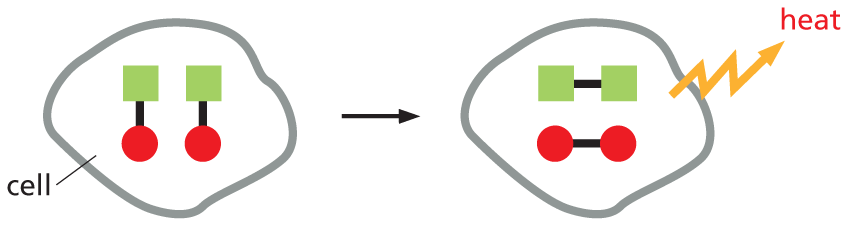
Think of a chemical reaction occurring in a cell that has a constant temperature and volume. This reaction can produce disorder in two ways.
1Changes in bond energy of the reacting molecules can cause heat to be released, which disorders the environment around the cell.
More information
Panel titled reactions cause disorder An introductory text reads, Think of a chemical reaction occurring in a cell that has a constant temperature and volume. This reaction can produce disorder in two ways.
1. Changes of bond energy of the reacting molecules can cause heat to be released, which disorders the environment around the cell.
An illustration shows a cell has two different molecules represented by a square bonded to a circle. This molecule is converted to another molecule in which the squares are paired and the circles are paired separately and heat is released.
2The reaction can decrease the amount of order in the cell—for example, by breaking apart a long chain of molecules, or by disrupting an interaction that prevents bond rotations.
More information
2. The reaction can decrease the amount of order in the cell, for example, by breaking apart a long chain of molecules, or by disrupting an interaction that prevents bond rotations.
An illustration shows molecules in a cell arranged to form a chain. This cell is converted to another cell, in which the molecules are separated.
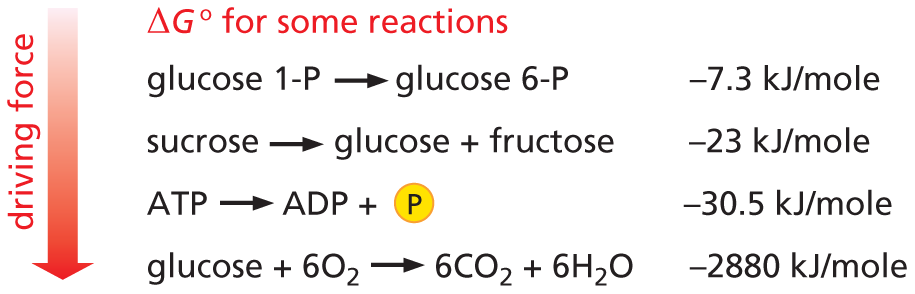
PREDICTING REACTIONS
To predict the outcome of a reaction (Will it proceed to the right or to the left? At what point will it stop?), we must determine its standard free-energy change (ΔG°). This quantity represents the gain or loss of free energy as 1 mole of reactant is converted to 1 mole of product under “standard conditions” (all molecules present in aqueous solution at a concentration of 1 M and pH 7.0).
More information
Panel titled predicting reactions. An introductory text reads, to predict the outcome of a reaction (Will it proceed to the right or to the left? At what point will it stop?), we must determine its standard free-energy change (delta G degree). This quantity represents the gain or loss of free energy as one mole of reactant is converted to one mole of product under “standard conditions” (all molecules present in aqueous solution at a concentration of 1 M and p H 7.0).
Four reactions are shown as follows:
Glucose 1-P yields glucose 6-P; Delta G of this reaction equals negative 7.3 Kilojoules per mole.
Sucrose yields glucose and fructose; Delta G of this reaction equals negative 23 Kilojoules per mole.
A T P yields A D P and phosphate; Delta G of this reaction equals negative 30.5 Kilojoules per mole.
Glucose combines with 6 O subscript 2 to form 6 C O subscript 2 + 6 H subscript 2 O; Delta G of this reaction equals negative 2867 Kilojoules per mole.
A downward arrow alongside indicates that the driving force of these reactions decreases from 1 through 4.
ΔG (“DELTA G”)
Changes in free energy occurring in a reaction are denoted by ΔG, where “Δ” indicates a difference. Thus, for the reaction
A + B ⟶ C + D
ΔG = free energy (C + D) minus free energy (A + B)
ΔG measures the amount of disorder caused by a reaction: the change in order inside the cell, plus the change in order of the surroundings caused by the heat released.
ΔG is useful because it measures how far away from equilibrium a reaction is. The reaction
More information
Panel titled delta G.
An introductory text reads, Changes in free energy occurring in a reaction are denoted by delta G, where “delta” indicates a difference. Thus, for the reaction A plus B yields C plus D.
Delta G equals free energy (C plus D) minus free energy (A plus B).
Text reads, Delta G measures the amount of disorder caused by a reaction: the change in order inside the cell, plus the change in order of the surroundings caused by the heat released.
Delta G is useful because it measures how far away from equilibrium a reaction is. The reaction: A T P yields A D P plus P has a large negative delta G because cells keep the reaction a long way from equilibrium by continually making fresh A T P. However, if the cell dies, then most of its A T P will be hydrolyzed until equilibrium is reached; at equilibrium, the forward and backward reactions occur at equal rates and delta G equals 0.
has a large negative ΔG because cells keep the reaction a long way from equilibrium by continually making fresh ATP. However, if the cell dies, then most of its ATP will be hydrolyzed until equilibrium is reached; at equilibrium, the forward and backward reactions occur at equal rates and ΔG = 0.
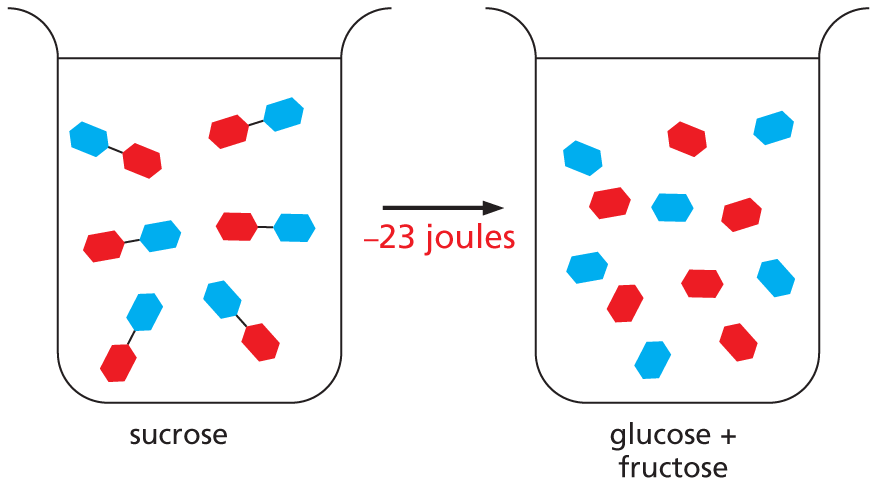
SPONTANEOUS REACTIONS
From the second law of thermodynamics, we know that the disorder of the universe can only increase. ΔG is negative if the disorder of the universe (reaction plus surroundings) increases.
In other words, a chemical reaction that occurs spontaneously must have a negative ΔG:
Gproducts − Greactants = ΔG < 0
EXAMPLE: The difference in free energy of 100 mL of 10 mM sucrose (common sugar) and 100 mL of 10 mM glucose plus 10 mM fructose is about –23 joules. Therefore, the hydrolysis reaction that produces two monosaccharides from a disaccharide (sucrose → glucose + fructose) can proceed spontaneously.
More information
Panel titled spontaneous reactions. An introductory text reads, from the second law of thermodynamics, we know that the disorder of the universe can only increase. Delta G is negative if the disorder of the universe (reaction plus surroundings) increases. In other words, a chemical reaction that occurs spontaneously must have a negative delta G:
G subscript products minus G subscript reactants equals Delta G less than 0.
Example: The difference in free energy of 100 m L of 10 m M sucrose (common sugar) and 100 m L of 10 m M glucose plus 10 m M fructose is about negative 23 joules. Therefore, the hydrolysis reaction that produces two monosaccharaides from a disaccharide (sucrose reacts to yield glucose and fructose) can proceed spontaneously.
An illustration shows a beaker containing sucrose molecules; each molecule has two units bound to each other. These molecules undergo hydrolysis and the molecules are broken down into glucose and fructose. 23 joules of heat is lost.
Text below reads, In contrast, the reverse reaction (glucose combines with fructose yields sucrose), which has a delta G of 23 joules, could not occur without an input of energy from a coupled reaction.
In contrast, the reverse reaction (glucose + fructose → sucrose), which has a ΔG of +23 joules, could not occur without an input of energy from a coupled reaction.
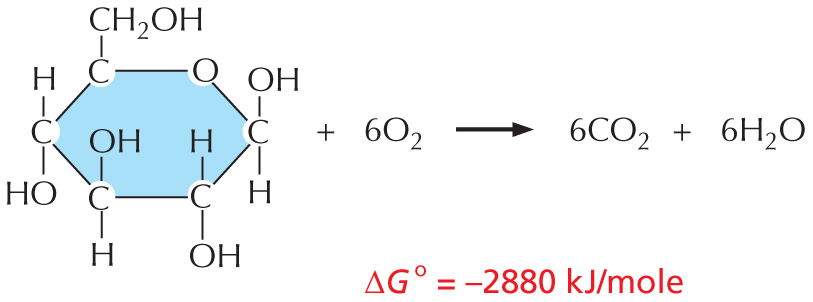
REACTION RATES
A spontaneous reaction is not necessarily a rapid reaction: a reaction with a negative free-energy change (ΔG) will not necessarily occur rapidly by itself. Consider, for example, the combustion of glucose in oxygen:
More information
Panel titled reaction rates. An introductory text reads, a spontaneous reaction is not necessarily a rapid reaction: a reaction with a negative free-energy change (delta G) will not necessarily occur rapidly by itself. Consider, for example, the combustion of glucose in oxygen:
Glucose combines with 6 O subscript 2 to yield 6 C O subscript 2 and 6 H subscript 2 O. Delta G of this reaction equals negative 2867 Kilojoules per mole.
The molecular structure of glucose has the following configuration: C 1, C 2, C 3, and C 4 are each bonded to a hydrogen and a hydroxyl group. C 5 is bonded to a C H subscript 2 O H group.
Text reads, even this highly favorable reaction may not occur for centuries unless enzymes are present to speed up the process. Enzymes are able to catalyze reactions and speed up their rate, but they cannot change the delta G degree of a reaction.
Even this highly favorable reaction may not occur for centuries unless enzymes are present to speed up the process. Enzymes are able to catalyze reactions and speed up their rate, but they cannot change the ΔG° of a reaction.
ΔG EQUALS ZERO AT CHEMICAL EQUILIBRIUM
There is a fixed relationship between the standard free-energy change of a reaction, ΔG°, and its equilibrium constant K. For example, the reversible reaction
More information
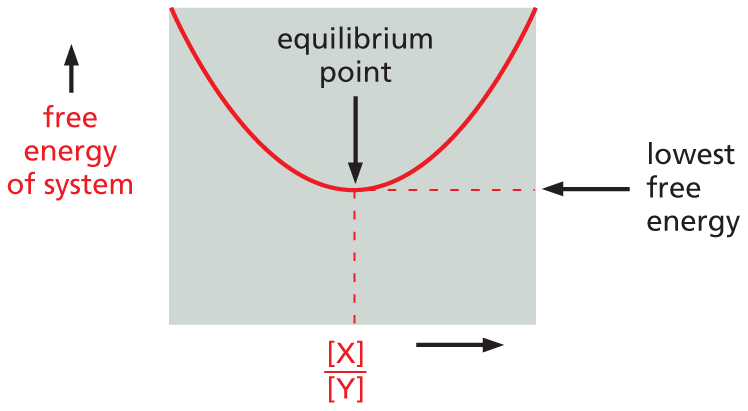
Panel titled delta G equals zero at chemical equilibrium. An introductory text reads, There is a fixed relationship between the standard free-energy change of a reaction, Delta G knot, and its equilibrium constant K. For example, the reversible reaction in which Y is converted to X, will proceed until the ratio of concentrations of X is to Y is equal to K. At this point, the free energy of the system will have its lowest value.
will proceed until the ratio of concentrations [X]/[Y] is equal to K (note: square brackets [ ] indicate concentration). At this point, the free energy of the system will be at its lowest value, and the reaction’s ΔG will be 0.
More information
A graph plotting free energy of system on the vertical axis against the concentration ratios of X to Y along the horizontal axis has a U shaped curve. Two dashed lines connect the equilibrium point of the curve to the two axes. The point where the dashed line meets the vertical axis represents lowest free energy.
At 37 degree Celsius,
Delta G equals 0 and the constant R T equals 2.58 so delta G knot equals negative 2.58 in K or negative 5.94 log K to the base 10. Therefore, K equals 10 to the power negative delta G knot over 5.94. A note alongside reads, see text, page 102
At 37°C, where ΔG = 0 and the constant RT = 2.58,
ΔG° = –2.58 In K
or, in the more familiar base-10 logarithm,
ΔG°= –5.94 logK
Therefore, K = 10−ΔG°/5.94 (see text, p. 102)
This equation allows the equilibrium constant, K, to be calculated from a reaction’s ΔG°. For example, the reaction
More information
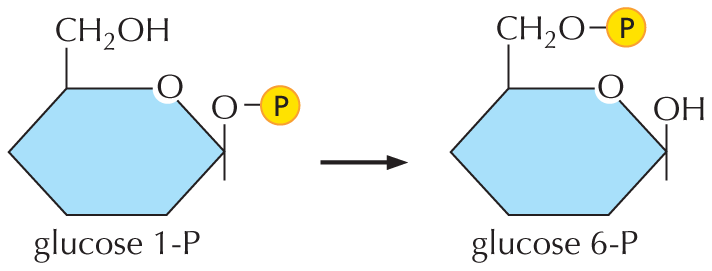
For example, the reaction:
Glucose 1-P yielding glucose 6-P, Delta G 0 equals negative 7.3 Kilojoules per mole. Therefore, its equilibrium constant K = 10 to the power of (7.3 over 5.94) equals 10 to the power 1.23 equals 17.
So the reaction will reach steady state when the concentration of glucose 6-P over the concentration of glucose 1-P equals 17.
The molecular structure of Glucose 1-P shows an oxygen substituted five membered ring in which C 1 is bonded to an oxygen atom which is further bonded to a phosphate group; C 5 is bonded to a C H subscript 2 O H. The structure of glucose 6-P shows an oxygen substituted five membered ring in which C 1 is bonded to a hydroxyl group and C 5 is single bonded to a C H subscript 2 O group which is further bonded to a phosphate group.
has ΔG° = –7.3 kJ/mole. Therefore, its equilibrium constant
K = 10(7.3/5.94) = 10(1.23) = 17
So the reaction will reach steady state when
[glucose 6-P]/[glucose 1-P] = 17
COUPLED REACTIONS
Reactions can be “coupled” together if they share one or more intermediates. In this case, the overall free-energy change is simply the sum of the individual ΔG° values. A reaction that is unfavorable (has a positive ΔG°) can for this reason be driven by a second, highly favorable reaction.
SINGLE REACTION
More information
Panel titled coupled reactions. An introductory text reads, Reactions can be “coupled” together if they share one or more intermediates. In this case, the overall free-energy change is simply the sum of the individual delta G degree values. A reaction that is unfavorable (has a positive delta G degree) can for this reason be driven by a second, highly favorable reaction.
Single reaction:
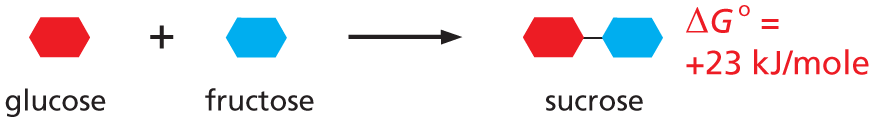
Glucose (represented by a red ring) combines with fructose (represented by a blue ring) reacts to yield sucrose (represented by a red ring bonded to a blue ring), where delta G degree = 23 Kilojoules per mole.
Net result: reaction will not occur.
NET RESULT:reaction will not occur
More information
A T P reacts to yield A D P and phosphate, delta G 0 of this reaction equals negative 30.5 kilojoules per mole.
Net result: reaction is highly favorable.
Coupled reactions:
NET RESULT:reaction is highly favorable
COUPLED REACTIONS
More information
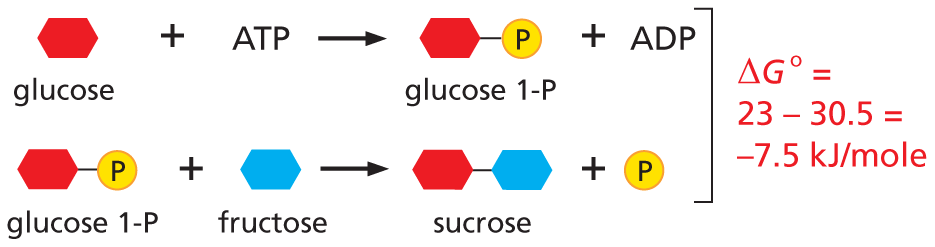
Glucose (represented by a red cyclohexane) combines with A T P to yield glucose 1-P (represented by a red ring bonded to phosphate) plus A D P.
Glucose 1-P (represented by a red ring bonded to phosphate) combines with fructose (represented by a blue ring) yields sucrose (represented by a red ring bonded to blue ring) plus a phosphate. Delta G 0 equals 23 minus 30.5 equals negative 7.5 kilojoules per mole.
Net result: sucrose is made in a reaction driven by the hydrolysis of A T P.
NET RESULT:sucrose is made in a reaction driven by the hydrolysis of ATP
“HIGH-ENERGY” BONDS
One of the most common reactions in the cell is hydrolysis, in which a covalent bond is split by adding water.
More information
Panel titled high energy bonds An introductory text reads, one of the most common reactions in the cell is hydrolysis, in which a covalent bond is split by adding water.
Molecule A bonded to molecule B undergoes hydrolysis to yield molecule A bonded to a hydroxyl group plus a hydrogen atom bonded to a molecule B.
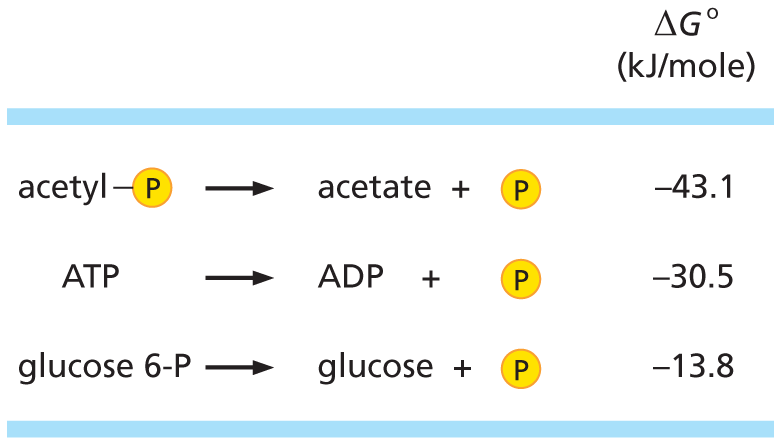
The delta G naught for this reaction is sometimes loosely termed the bond energy. Compounds such as acetyl phosphate and A T P, which have a large negative delta G degree of hydrolysis in an aqueous solution, are said to have “high-energy” bonds.
The ΔG° for this reaction is sometimes loosely referred to as “bond energy.” Compounds such as acetyl phosphate and ATP are said to have “high-energy” bonds because these compounds have a large negative ΔG° of hydrolysis in an aqueous solution.
More information
Acetyl bonded with phosphate reacts to yield acetate and phosphate, delta G naught equals negative 43.1 Kilojoules per mole.
A T P reacts to yield A D P and phosphate, delta G naught equals negative 30.5 Kilojoules per mole.
Glucose 6-P reacts to yield glucose and phosphate, delta G naught equals negative 13.8 Kilojoules per mole.
(Note that, for simplicity, water is omitted from the above equations.)
(Note that, for simplicity, H2O is omitted from the above equations.)
For Sequential Reactions, the Changes in Free Energy Are Additive
Now we return to our original concern regarding how cells can generate and maintain order. More specifically: how can enzymes catalyze reactions that are energetically unfavorable?
One way they do so is by directly connecting energetically unfavorable reactions to energetically favorable ones. Consider, for example, two sequential reactions,
X ⟶ Y and Y ⟶ Z
where the ΔG° values are +21 and –54 kJ/mole, respectively. (Recall that a mole is 6 × 1023 molecules of a substance.) The unfavorable reaction, X ⟶ Y, will not occur spontaneously. However, it can be driven by the favorable reaction Y ⟶ Z, provided that the second reaction follows the first. That’s because the overall free-energy change for the two reactions is equal to the sum of the free-energy changes for each individual reaction. In this case, the ΔG° for the coupled reaction, X ⟶ Y ⟶ Z, will be –33 kJ/mole, making the overall pathway energetically favorable.
QUESTION 3–5
For the reactions shown in Figure 3–20, sketch an energy diagram similar to that in Figure 3–12 for the two reactions alone and for the combined reactions. Indicate the standard free-energy changes for the reactions X ⟶ Y, Y ⟶ Z, and X ⟶ Z in the drawing. Show how enzymes that catalyze these reactions would change the energy diagram.
Cells can therefore cause the energetically unfavorable transition, X ⟶ Y, to occur if an enzyme catalyzing the X ⟶ Y reaction is supplemented by a second enzyme that catalyzes the energetically favorable reaction, Y ⟶ Z. In effect, the reaction Y ⟶ Z acts as a “siphon,” pulling the conversion of all of molecule X to molecule Y, and then to molecule Z (Figure 3–20). Several of the reactions in the long pathway that converts sugars such as glucose into CO2 and H2O are energetically unfavorable. This life-giving process nevertheless proceeds rapidly to completion because the total ΔG° for the series of sequential reactions has such a large negative value: –2880 kJ/mole.
More information
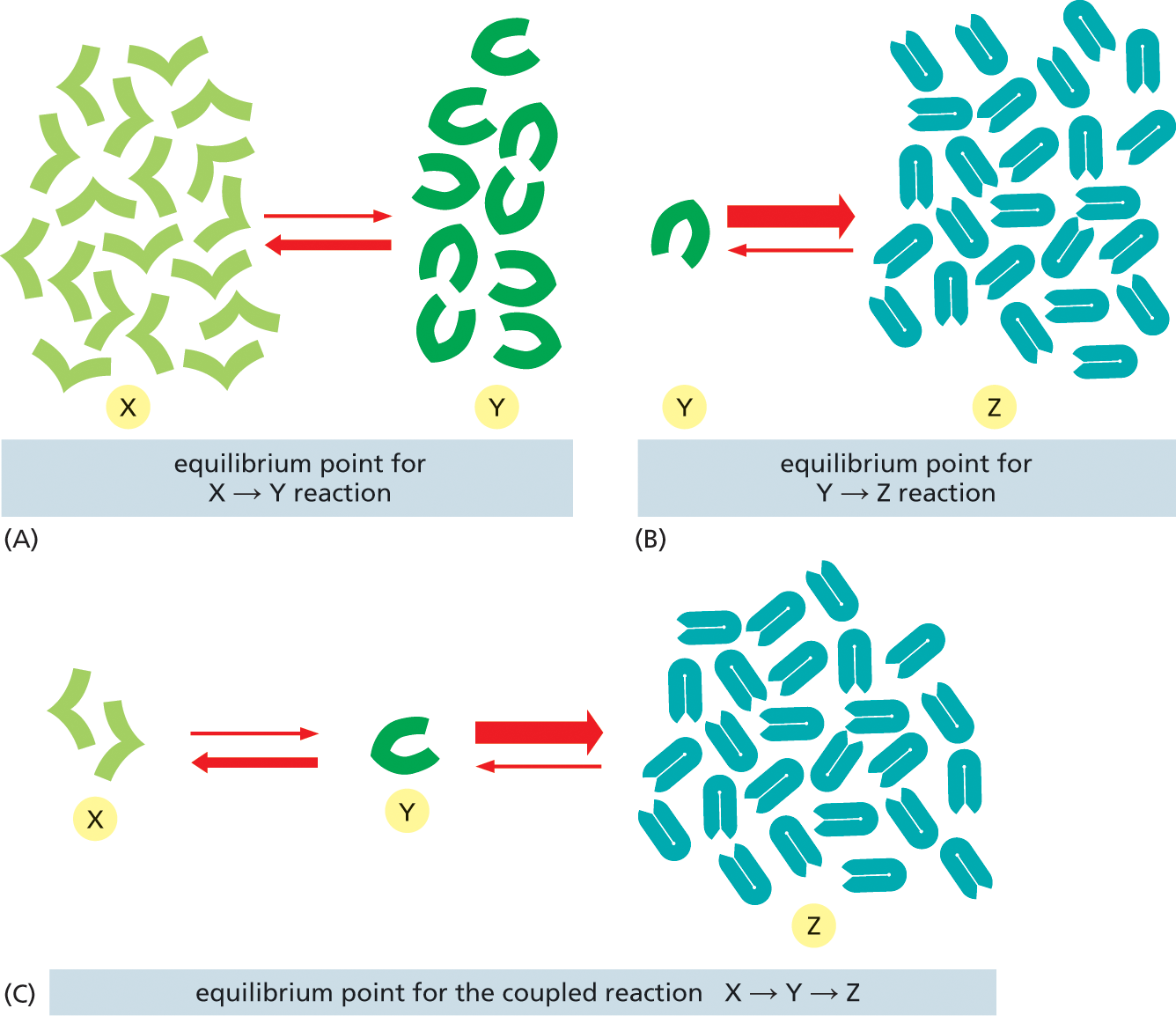
A set of three illustrations depicts how the equilibria of sequential reactions can have the net effect of a chemical siphon. The illustration (A) titled “equilibrium point for X converted into Y reaction” shows a cluster of X molecules converted into Y molecules. The conversion of X to Y has a lower rate of conversion; whereas the conversion of Y to X has a higher rate of conversion.
The illustration (B) titled “equilibrium point for Y converted into Z reaction” shows a Y molecule which is converted into a cluster of Z molecules; this forward reaction has a high rate of conversion and the reverse reaction has a lesser rate of conversion.
The illustration (C) titled “equilibrium point for the coupled reaction X converted into Y which is converted into Z” shows an X molecule converted into a Y molecule and the Y molecule is further converted into a cluster of Z molecules. X to Y has a lesser rate of conversion; Y to X has a higher rate of conversion; Y to Z has a higher rate of conversion and Z to Y has a lesser rate of conversion.
Figure 3–20An energetically unfavorable reaction can be driven by an energetically favorable follow-on reaction that acts as a chemical siphon. (A) At equilibrium, there are twice as many X molecules as Y molecules. (B) At equilibrium, there are 25 times more Z molecules than Y molecules. (C) If the reactions in (A) and (B) are coupled, nearly all of the X molecules will be converted to Z molecules, as shown. In terms of energetics, the ΔG° of the Y ⟶ Z reaction is so negative that, when coupled to the X ⟶ Y reaction, it lowers the ΔG of X ⟶ Y. This is because the ΔG of the X ⟶ Y reaction decreases as the ratio of Y to X decreases (see Figure 3–18).
Forming a sequential pathway, however, is not the answer for all of a cell's metabolic needs. In many cases, the desired is simply X ⟶ Y, without further conversion of Y to some other product. Fortunately, there are other, more general ways of using enzymes to couple reactions together, involving the production of activated carriers that can shuttle energy from one reaction site to another, as we discuss shortly.
Enzyme-catalyzed Reactions Depend on Rapid Molecular Collisions
For two molecules to interact, they must first collide—an event that depends on the concentrations of the participants (see Figure 3–19). However, enzymes and their substrates are generally found in relatively small quantities in the cell. The most abundant substrates, for example, are present at a concentration of about 0.5 mM. Because pure water is 55 M, that means that only one substrate molecule will be present in every 105 water molecules. Yet a typical enzyme can capture and process about 1000 substrate molecules every second—releasing its product and binding a new substrate in a fraction of a millisecond. How, then, do all of these molecules manage to find each other so quickly?
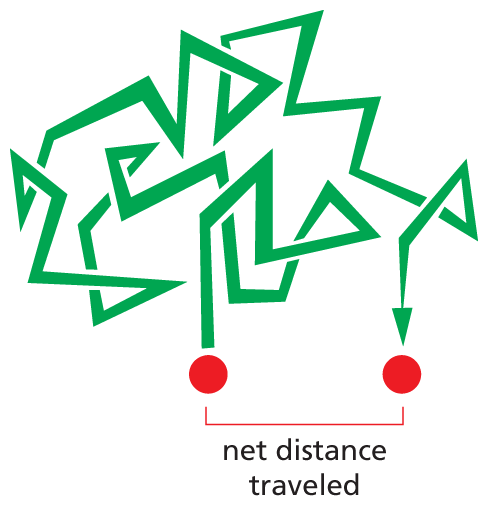
Inside cells, rapid chemical encounters are possible because molecular motions are enormously fast—very much faster than the human mind can easily imagine. Because of heat energy, molecules are in constant rapid motion and consequently will explore the cytosolic space very efficiently by wandering randomly through it—a process called diffusion. In this way, every molecule in the cytosol collides with a huge number of other molecules each second. As these molecules in solution collide and bounce off one another, an individual molecule moves first one way and then another, its path constituting a random walk (Figure 3–21).
More information
An illustration shows the random walk of a molecule. The molecule moves in a variety of directions before reaching the end location. The net distance traveled between the starting and end location is much shorter than the distance traveled on the walk.
Figure 3–21A molecule traverses the cytosol by taking a random walk. Molecules in solution move in a random fashion due to the continual buffeting they receive in collisions with other molecules. This movement allows small molecules to diffuse rapidly throughout the cell cytosol (Movie 3.2). (Dynamic Figure) Although a molecule moving randomly by diffusion (red) might not wind up far from where it started (pink), it can explore an extensive region (green) of the cytosolic space.
Of course, the cytosol of a cell is densely packed with molecules of various shapes and sizes (Figure 3–22). Nevertheless, experiments in which fluorescent dyes and other labeled molecules are injected into the cell cytosol show that small organic molecules diffuse through this aqueous gel nearly as rapidly as they do through water. A small organic molecule, such as a substrate, takes only about one-fifth of a second on average to diffuse a distance of 10 μm.
More information
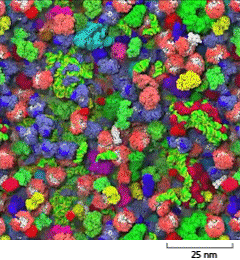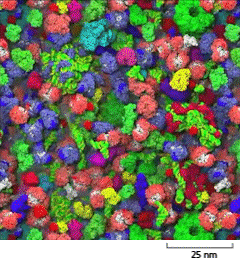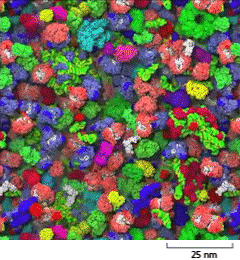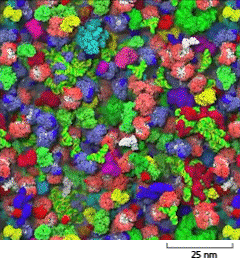
A schematic shows closely packed macromolecules in the cytosol on a scale of 25 nanometers. The macromolecules are lumpy and each is about 6 nanometers wide.
Figure 3–22The cytosol is crowded with large molecules. Only the macromolecules, which are drawn to scale and displayed in different colors, are shown. Enzymes and other macromolecules diffuse relatively slowly in the cytosol, in part because they interact with so many other macromolecules. Small molecules, by contrast, can diffuse nearly as rapidly as they do in water (see Movie 1.2). [From S.R. McGuffee and A.H. Elcock, PLoS Comput. Biol. 6(3):e1000694, 2010.] (Dynamic Figure) Diffusion of the macromolecules is shown in the simulation below. (Courtesy of Adrian Elcock, University of Iowa Carver College of Medicine.)
Because proteins diffuse through the cytosol much more slowly than do small molecules, it is the substrate that finds its enzyme—rather than the other way around. For a substrate present at a concentration of 0.5 mM, the site on an enzyme that binds this substrate will be bombarded by about 500,000 random collisions every second! For a substrate concentration tenfold lower (0.05 mM), the number of collisions drops to 50,000 per second, and so on. These fantastically numerous collisions play a critical role in life’s chemistry.
Noncovalent Interactions Allow Enzymes to Bind Specific Molecules
The first step in any enzyme-catalyzed chemical reaction is the binding of the substrate. Once this step has taken place, the substrate must remain bound to the enzyme long enough for the chemistry to occur. The association of enzyme and substrate is stabilized by the formation of multiple, weak bonds between the participating molecules. These weak interactions—which can include hydrogen bonds, van der Waals attractions, and electrostatic attractions (discussed in Chapter 2)—persist until random thermal motion causes the molecules to dissociate again.
When two colliding molecules have poorly matching surfaces, few noncovalent bonds are formed, and their total energy is negligible compared with that of thermal motion. In this case, the two molecules dissociate as rapidly as they come together (see Figure 2–35). Poor noncovalent bond formation is what prevents unwanted associations from forming between mismatched molecules, such as those between an enzyme and the wrong substrate. Only when the enzyme and substrate are well matched do they form many weak interactions that keep them together long enough for a covalent bond in the substrate molecule to be formed or broken. The binding strength of an enzyme and substrate is a reflection of the equilibrium constant that governs their interactions, as we discuss next.
The Equilibrium Constant Reflects the Strength of Noncovalent Binding Interactions
Noncovalent interactions are immensely important to cells. They make possible not only the binding of substrates to enzymes, but also the binding of proteins to DNA or to other proteins—interactions needed to produce the many different structural and functional protein complexes required in a living cell.
Two molecules will bind to each other if the free energy of the resulting complex is lower than the sum of the free energies of the two partners when unbound. Because the equilibrium constant (K) is directly related to ΔG°, K is commonly used to measure the binding strength of a noncovalent interaction between two molecules. When molecule A binds to molecule B to form the complex AB, the reaction proceeds until it reaches equilibrium. At this point, the concentrations of reactants A and B, and of the complex AB, can be used to determine the equilibrium constant K, and thus the ΔG° for that binding interaction (see Figure 3–19).
The equilibrium constant for binding, denoted as K = [AB]/[A][B], is known as the affinity constant. It becomes larger as the binding energy—that is, the free energy released when two molecules interact—increases. In other words, the larger K is, the greater is the drop in free energy between the dissociated and associated states, and the more tightly the two molecules will bind. Even a change of a few noncovalent bonds can have a striking effect on a binding interaction. Thus, in the example illustrated in Figure 3–23, a loss of a few hydrogen bonds from a binding interaction can be seen to cause a dramatic decrease in the amount of a complex at equilibrium.
More information
A textbox shows an example of the presence and absence of cytosolic proteins in weak noncovalent bonds. Text reads as follows: Consider 1000 molecules of A and 1000 molecules of B in the cytosol of a eukaryotic cell. The concentration of both will be about 10 to the power negative 9 M. If the equilibrium constant (K) for A plus B leads to A B is 10 to the tenth power liters per mole, then at equilibrium there will be 270 molecules of A, 270 molecules of B and 730 Complexes of A B.
If the equilibrium constant is a little weaker, say 108 liters per mole—a value that represents a loss of 11.9 Kilojoules per mole of binding energy from the example above, or 2 to 3 fewer hydrogen bonds—then there will be 915 Molecules of A, 915 Molecules of B, and 85 complexes of A B.
Figure 3–23Small changes in the number of weak bonds can have drastic effects on a binding interaction. This example illustrates the dramatic effect of the presence or absence of a few weak noncovalent bonds in the interaction between two cytosolic proteins.
QUESTION 3–6
In cells, an enzyme catalyzes the reaction AB ⟶ A + B. It was isolated, however, as an enzyme that carries out the opposite reaction, A + B ⟶ AB. Explain the paradox.
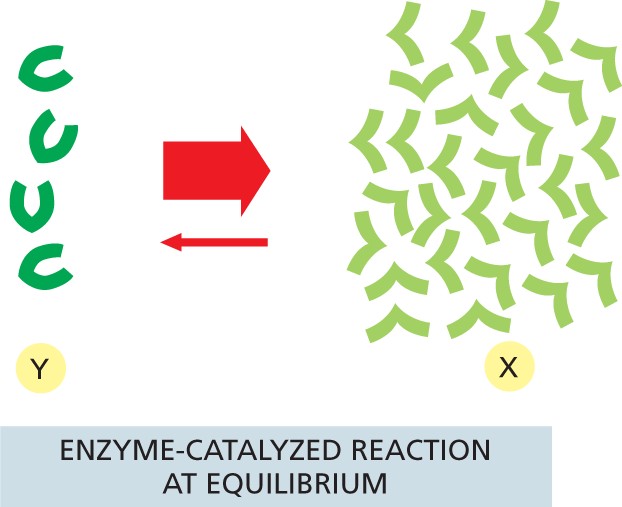
It is important to note that, because the same noncovalent bonds are formed between an enzyme and its substrate whether the reaction goes forward or backward, an enzyme that lowers the activation energy for the reaction Y ⟶ X will also lower the activation energy for the reverse reaction, X ⟶ Y, by exactly the same amount. This means that the forward and backward reactions are always accelerated by the same factor by an enzyme—a requirement given that the reaction’s ΔG° does not change (see Figure 3–12). Thus the equilibrium constant for the catalyzed reaction is the same as that of the uncatalyzed reaction (Figure 3–24).
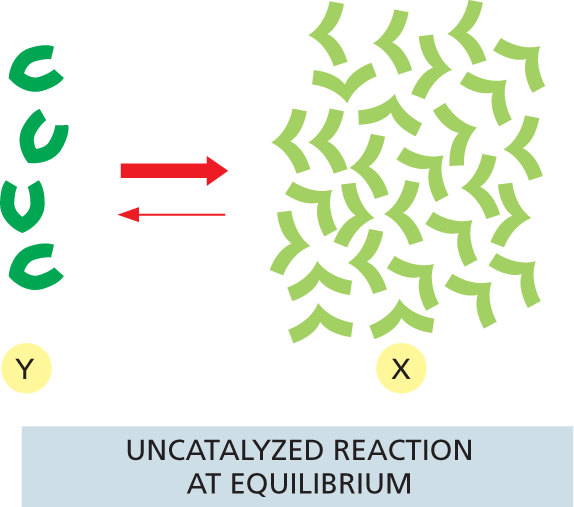
More information
An illustration shows an uncatalyzed reaction at equilibrium. The forward reaction has a moderate conversion rate whereas the reverse reaction has a low conversion rate.
(A)More information
An illustration shows an enzyme catalyzed reaction at equilibrium. The forward reaction has a high conversion rate whereas the reverse reaction has a low conversion rate.
(B)Figure 3–24Enzymes cannot change the equilibrium point for reactions. Enzymes, like all catalysts, speed up the forward and reverse rates of a reaction by the same amount. Therefore, for both the (A) uncatalyzed and (B) catalyzed reactions shown here, the number of molecules undergoing the transition Y ⟶ X is equal to the number of molecules undergoing the transition X ⟶ Y when the ratio of X molecules to Y molecules is 7 to 1, as illustrated. In other words, both the catalyzed and uncatalyzed reactions will eventually reach the same equilibrium point, although the catalyzed reaction will reach equilibrium much faster.
“Delta G”: in a chemical reaction, the difference in free energy between reactant and product molecules. A large negative value of ΔG indicates that the reaction has a strong tendency to occur. (See alsostandard free-energy change.)