Much of the energy released by an energetically favorable reaction, such as the oxidation of a food molecule, must be stored temporarily before it can be used by cells to fuel energetically unfavorable reactions, such as the synthesis of all the other molecules needed by the cell. In most cases, the energy is stored as chemical-bond energy in a set of activated carriers, small organic molecules that contain one or more energy-rich covalent bonds. These molecules diffuse rapidly and carry their bond energy from the sites of energy generation to the sites where energy is used either for biosynthesis or for the many other energy-requiring activities that a cell must perform (Figure 3–25). In a sense, cells use activated carriers like money to pay for the energetically unfavorable reactions that otherwise would not take place.
More information
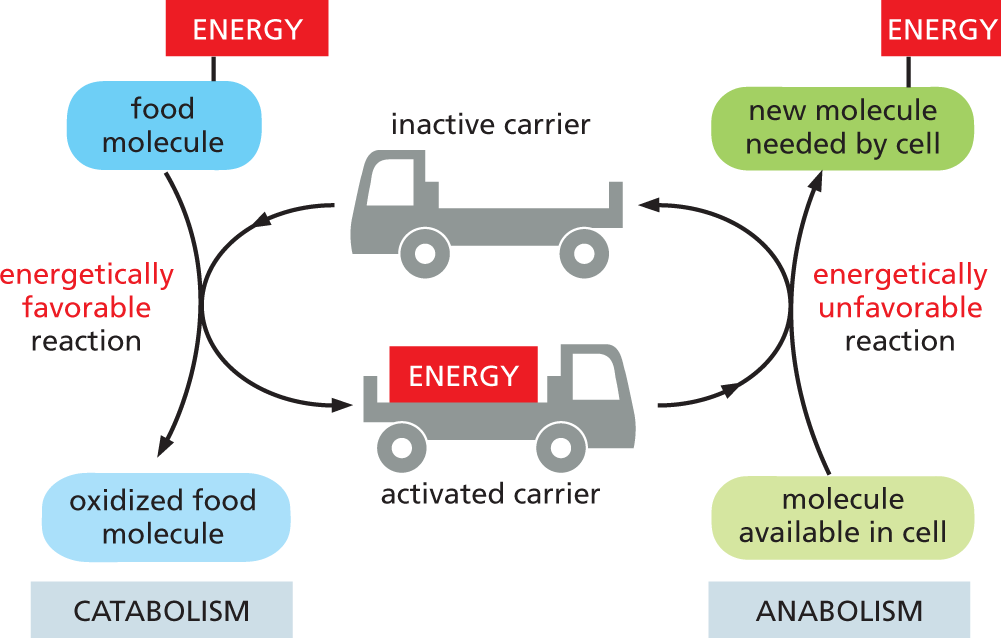
An illustration shows how carriers can store and transfer energy in metabolism. The chart is divided into two parts representing catabolism and anabolism. In catabolism, the energy from the food molecules is taken by the inactive carrier so the food molecules get oxidized to form activated carriers. This reaction is termed as an energetically favorable reaction. In anabolism, the activated carriers combine with the molecules in the cell to produce new molecules which are needed by the cell. This reaction is termed as an energetically unfavorable reaction.
Figure 3–25Activated carriers can store and transfer energy in a form that cells can use. By serving as intracellular energy shuttles, activated carriers perform their function as go-betweens that link the release of energy from the breakdown of food molecules (catabolism) to the energy-requiring biosynthesis of small and large organic molecules (anabolism).
Activated carriers store energy in an easily exchangeable form, either as a readily transferable chemical group or as readily transferable (“high-energy”) electrons. They can serve a dual role as a source of both energy and chemical groups for biosynthetic reactions. As we shall discuss shortly, the most important activated carriers are ATP and two molecules that are close chemical cousins, NADH and NADPH.
The Formation of an Activated Carrier Is Coupled to an Energetically Favorable Reaction
When a fuel molecule such as glucose is oxidized inside a cell, enzyme-catalyzed reactions ensure that a large part of the free energy released is captured in a chemically useful form, rather than being released wastefully as heat. When your cells oxidize the sugar from a chocolate bar, that energy allows you to power metabolic reactions; burning that same chocolate bar in the street will get you nowhere, warming the environment while producing no metabolically useful energy.
In cells, energy capture is achieved by means of a coupled reaction. Here, an energetically favorable reaction is used to drive an energetically unfavorable one, so that an activated carrier or some other useful molecule is produced. This coupling requires enzyme catalysis, and it is fundamental to all of the energy transactions in the cell.
More information
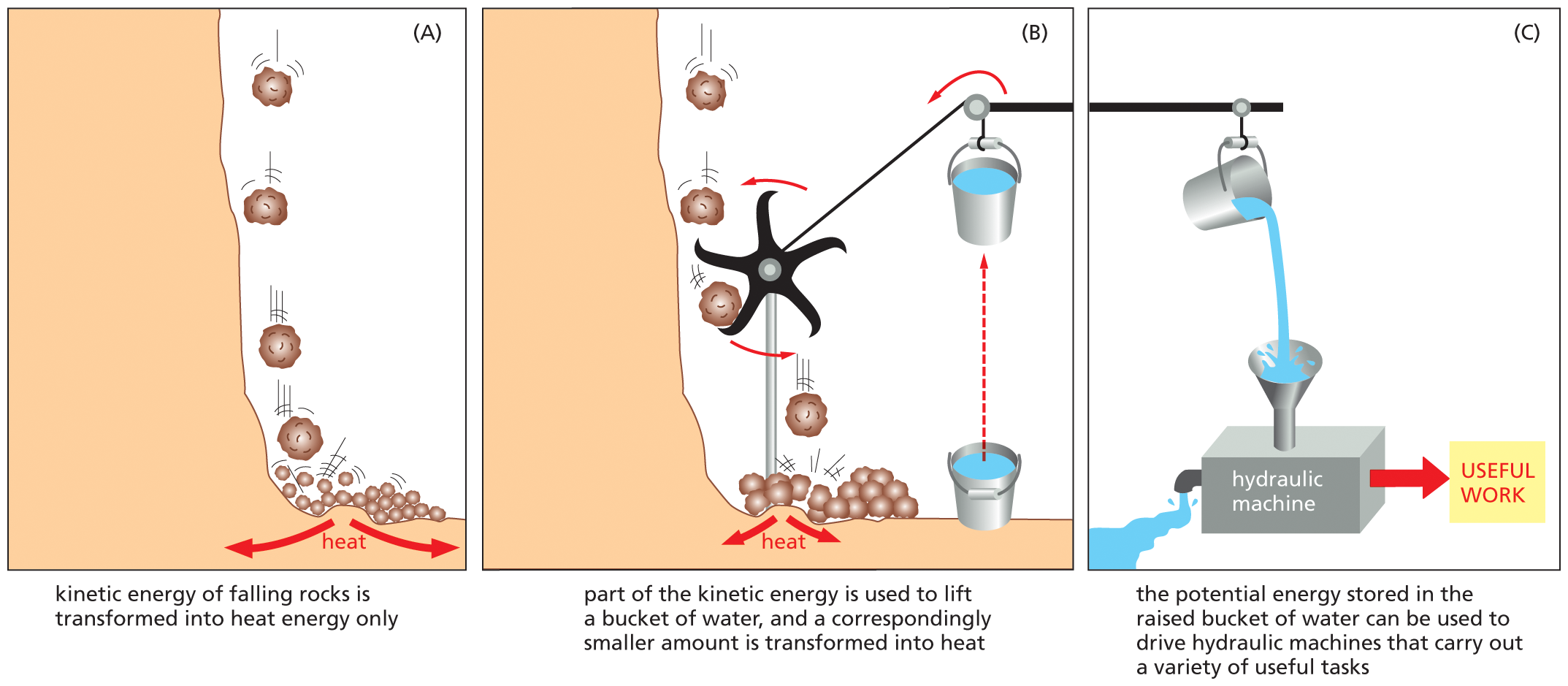
An illustrated analogy depicts the principle of coupled chemical reactions. Part A. A schematic shows rocks falling from a height. Heat energy is released in the process. Text reads, kinetic energy of falling rocks is transformed into heat energy only.
Part B. A schematic shows a turbine which is positioned in the path of the falling rocks. The blades of the turbine rotate in a counterclockwise direction. The turbine is attached to a pulley, which is tied to a bucket of water. Text reads, part of the kinetic energy is used to lift a bucket of water and a correspondingly smaller amount is transformed into heat. Part C. The bucket of water is raised to a height, and water from it spills into a funnel attached to a hydraulic machine, which performs useful work. Water flows out of the other side of the machine. Text reads, the potential energy stored in the raised bucket of water can be used to drive hydraulic machines that carry out a variety of useful tasks.
Figure 3–26A mechanical model illustrates the principle of coupled chemical reactions. (A) The spontaneous reaction shown could serve as an analogy for the direct oxidation of glucose to CO2 and H2O, which produces only heat. (B) The same reaction is coupled to a second reaction, which could serve as an analogy for the synthesis of activated carriers. (C) The energy captured in the raised bucket in (B) can be used to drive a variety of otherwise energetically unfavorable reactions.
QUESTION 3–7
Figure 3–26B illustrates how an energetically favorable reaction can provide energy to drive an energetically unfavorable reaction. Consider the energetically unfavorable conversion of molecule X to molecule Y, which is driven by the hydrolysis of ATP:
X + ATP ⟶ Y + ADP + phosphate
For this reaction, which molecule or molecules would be analogous to (i) the rocks at the top of the cliff, (ii) the broken debris at the bottom of the cliff, (iii) the bucket at its highest point, and (iv) the bucket on the ground?
What would be analogous to (i) the rocks hitting the ground in the absence of the paddle wheel in Figure 3–26B and (ii) the hydraulic machine in Figure 3–26C?
To provide an everyday representation of how coupled reactions work, let’s consider a mechanical analogy in which an energetically favorable chemical reaction is represented by rocks falling from a cliff. The kinetic energy of falling rocks would normally be entirely wasted in the form of heat generated by friction when the rocks hit the ground (Figure 3–26A). By careful design, however, part of this energy could be used to drive a paddle wheel that lifts a bucket of water (Figure 3–26B). Because the rocks can now reach the ground only after moving the paddle wheel, we say that the energetically favorable reaction of rocks falling has been directly coupled to the energetically unfavorable reaction of lifting the bucket of water. Because part of the energy is used to do work in (B), the rocks hit the ground with less velocity than in (A), and correspondingly less energy is wasted as heat. The energy saved in the elevated bucket of water can then be used to do useful work (Figure 3–26C).
Analogous processes occur in cells, where enzymes play the role of the paddle wheel in Figure 3–26B. By mechanisms that we discuss in Chapter 13, enzymes couple an energetically favorable reaction, such as the oxidation of food molecules, to an energetically unfavorable reaction, such as the generation of activated carriers. As a result, the amount of heat released by the oxidation reaction is reduced by exactly the amount of energy that is stored in the energy-rich covalent bonds of the activated carrier. That saved energy can then be used to power a chemical reaction elsewhere in the cell.
An understanding of how the energy locked in food molecules is converted into a form that is useful for the cell required the dedicated effort of the world’s finest chemists. Their discoveries, amassed over the first half of the twentieth century, marked the dawn of the study of biochemistry (How We Know, pp. 107–108).
HOW WE KNOW
“HIGH-ENERGY” PHOSPHATE BONDS POWER CELL PROCESSES
Cells require a continuous stream of energy to generate and maintain order, while acquiring the materials they need to survive, grow, and reproduce. But even as late as 1921, very little was known about how energy—which for animal cells is derived from the breakdown of nutrients—is biochemically transformed, stored, and released for work in the cell. It would take the efforts of a handful of biochemists, many of whom worked with Otto Meyerhof—a pioneer in the field of cell metabolism—to get a handle on this fundamental problem.
Muscling in
Meyerhof was trained as a physician in Heidelberg, Germany, and he had a strong interest in physiological chemistry; in particular, he wondered how energy is transformed during chemical reactions in cells. He recognized that between its initial entry in the form of food and its final dissipation as heat, a large amount of energy must be made available by a series of intermediate chemical steps that allow the cell or organism to maintain itself in a state of dynamic equilibrium.
To explore how these mysterious chemical transformations power the work done by cells, Meyerhof focused his attention on muscle. Muscle tissue could be isolated from an animal, such as a frog, and stimulated to contract with a pulse of electricity. This muscle twitching provided a dramatic demonstration of the conversion of energy to a usable, mechanical form.
When Meyerhof got started, all that was known about the chemistry of contraction is that, in active muscle tissue, oxygen becomes depleted and lactic acid builds up. As Meyerhof’s first order of business, he demonstrated that this lactic acid comes from the breakdown of glycogen—a branched polymer made of glucose units that serves as an energy store in animal cells, particularly in muscle (see Panel 2–4, p. 77).
While Meyerhof focused on the chemistry, English physiologist Archibald “A.V.” Hill determined that working muscles give off heat, both as they contract and as they recover; further, he found that the amount of heat correlates with how hard the muscle is working.
Hill and Meyerhof then showed that the heat produced during muscle relaxation was linked to the resynthesis of glycogen. During the recovery period that follows a series of muscle contractions—when oxygen levels are restored—a portion of the lactic acid made during muscle contraction would be completely oxidized to CO2 and water; and the energy released by this oxidative breakdown would then be used to convert the remaining lactic acid back to glycogen (Figure 3–27). The conversion of glycogen to lactic acid—and back again—provided the first evidence of cyclical energy transformation in cells; it also earned Meyerhof and Hill a 1922 Nobel Prize.
More information
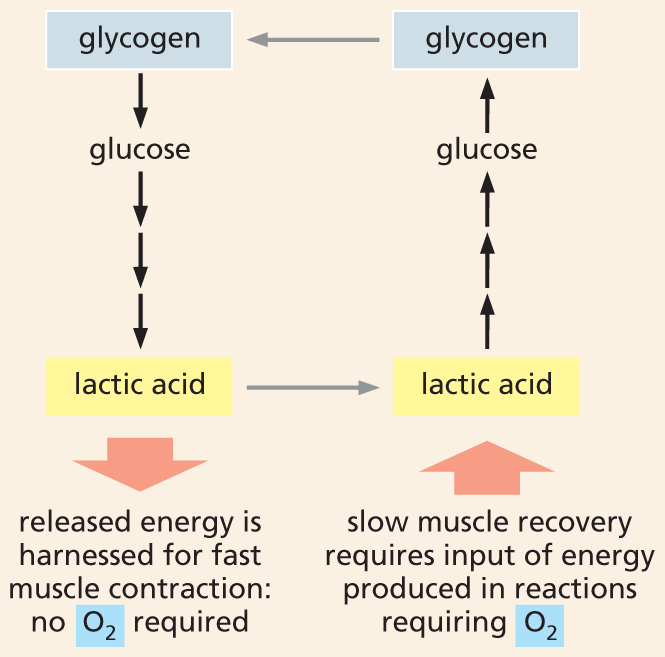
A flow chart shows the formerly hypothesized lactic acid cycle. Glycogen breaks down into glucose which forms lactic acid which releases energy. The released energy is harnessed for fast muscle contraction; no oxygen is required. Slow muscle recovery requires input of energy produced in the reactions requiring oxygen, which converts lactic acid into glucose; glucose is further converted into glycogen and thus the cycle continues.
Figure 3–27A “lactic acid cycle” was thought to supply the energy needed to power muscle contraction. Preparations of frog muscle were stimulated to contract. This rapid contraction, which depleted the muscle’s oxygen reserves, was accompanied by the breakdown of glycogen and the formation of lactic acid. The energy released by this glycogen breakdown was thought to somehow power muscle contraction. As the muscle slowly recovers, the replenished oxygen allowed the oxidative breakdown of a portion of the lactic acid to CO2 and H2O. The energy released by this lactic acid breakdown powered the conversion of the remaining lactic acid back to glycogen. Heat is generated during both the contraction and recovery phase.
In the mail
But did the conversion of glycogen into lactic acid directly power the mechanical work of muscle contraction? Meyerhof had thought so—until 1927, when a letter arrived from Danish physiologist Einar Lundsgaard. In it, Lundsgaard told Meyerhof of the surprising results of some experiments he had performed both on isolated muscles and in living rabbits and frogs. Lundsgaard had injected muscles with iodoacetate, a compound that inhibits an enzyme involved in the breakdown of sugars (discussed in Chapter 13). In these iodoacetate-treated muscles, fermentation was blocked and no lactic acid could be made.
What Lundsgaard discovered was that the poisoned muscles continued to contract. Indeed, animals injected with the compound at first “behaved quite normally,” wrote Fritz Lipmann, a biochemist who was working in Meyerhof’s laboratory. But after a few minutes, they suddenly keeled over, their muscles frozen in rigor.
But if the oxidation of lactic acid was not providing fuel for muscle contraction, what was? Lundsgaard went on to show that the source of energy for muscle contraction in poisoned muscles appeared to be a recently discovered molecule called creatine phosphate. When lactic acid formation was blocked by iodoacetate, muscle contraction was accompanied by the hydrolysis of creatine phosphate. When the supply of creatine phosphate was exhausted, muscles seized up permanently.
This finding created quite a stir in the Meyerhof lab, as it contradicted Meyerhof’s theory that lactic acid formation powered muscle contraction. And it pointed toward not just an alternative molecule, but a whole new idea: that certain phosphate bonds, when hydrolyzed, could provide energy. “Lundsgaard had discovered that the muscle machine can be driven by phosphate-bond energy, and he shrewdly realized that this type of energy was ‘nearer,’ as he expressed it, to the conversion of metabolic energy into mechanical energy than lactic acid,” wrote Lipmann.
But rather than being upset, Meyerhof welcomed Lundsgaard to his lab in Heidelberg, where he was serving as director of the Kaiser Wilhelm Physiology Institute. There, Lundsgaard made very careful measurements showing that the breakdown of creatine phosphate—and the heat it generated—closely tracked the amount of tension generated by intact muscle.
The most direct conclusion that could be drawn from these observations is that the hydrolysis of creatine phosphate supplied the energy that powers muscle contraction. But in one of his papers published in 1930, Lundsgaard was careful to note that there was another possibility: that in normal muscle, both lactic acid formation and creatine phosphate hydrolysis transferred energy to a third, yet-to-be identified system. This is where ATP comes in.
Squiggle P
Even before Lundsgaard’s eye-opening observations, Meyerhof had an interest in the amount of energy contained in various metabolic compounds, particularly those that contained phosphate. He thought that metabolic energy sources might be identified by finding naturally occurring molecules that release unusually large amounts of heat when hydrolyzed. Creatine phosphate was one of those compounds. Another was ATP, which had been discovered in 1929—by Meyerhof’s assistant, chemist Karl Lohmann, and, at the same time, by biochemists Cyrus Fiske and Yellapragada Subbarow working in America.
By 1935, Lohmann had demonstrated that the hydrolytic breakdown of creatine phosphate occurs through the transfer of its phosphate group to ADP to form ATP. It is the hydrolysis of ATP that serves as the direct source of energy for muscle contraction; creatine phosphate provides a reservoir of “high-energy” phosphate groups that replenish depleted ATP and maintain the needed ratio of ATP to ADP (Figure 3–28).
More information
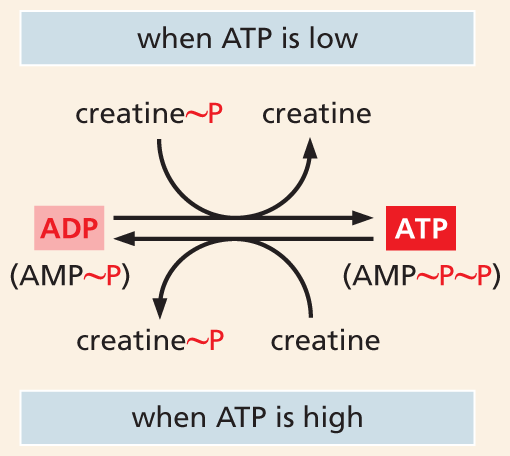
A flow chart shows how creatine phosphate stores energy. The chart shows that when A T P is low, creatine phosphate becomes creatine as A D P is converted to A T P. When A T P is high, creatine becomes creatine phosphate as A T P is converted to A D P. A D P is A M P plus one phosphate. A T P is A M P plus two phosphates.
Figure 3–28Creatine phosphate serves as an intermediate energy store. An enzyme called creatine kinase transfers a phosphate group from creatine phosphate to ADP when ATP concentrations are low; the same enzyme can catalyze the reverse reaction to generate a pool of creatine phosphate when ATP concentrations are high. Here, the “high-energy” phosphate bonds are symbolized by ~P. AMP is adenosine monophosphate (see Figure 3–41).
In 1941, Lipmann published a 63-page review in the inaugural issue of Advances in Enzymology entitled “The Metabolic Generation and Utilization of Phosphate Bond Energy.” This article introduced the symbol ~P (or “squiggle P”) to denote an energy-rich phosphate bond—one whose hydrolysis yields enough energy to drive energetically unfavorable reactions and processes (Figure 3–29).
More information
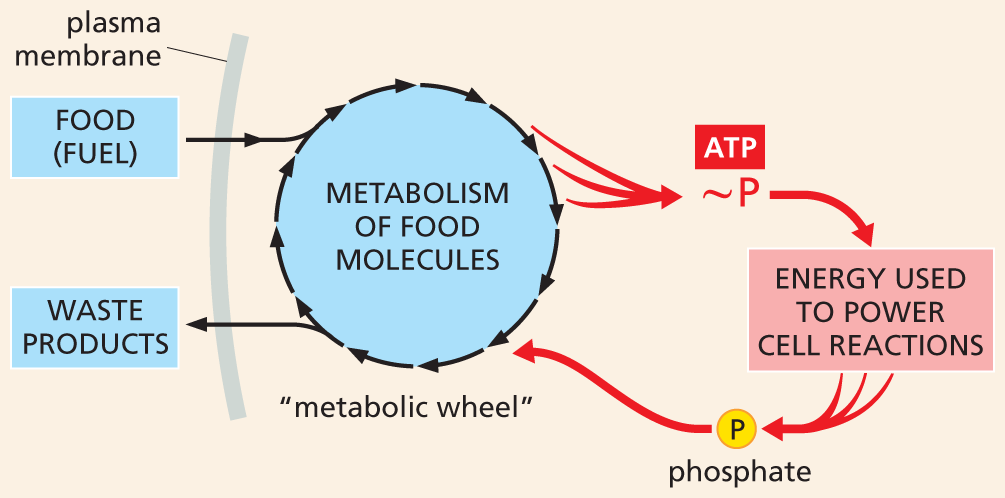
A flow chart depicts high energy phosphate bonds used to power cell reactions. The chart shows the food (fuel) entering through the plasma membrane followed by metabolism of food molecules in the metabolic wheel, which generates energy. This energy is captured by phosphate bond in A T P which powers cell reaction. The phosphate released is then transferred to the metabolic wheel and is eliminated out of the plasma membrane as waste products.
Figure 3–29High-energy phosphate bonds generate an energy current (red) that powers cell reactions. This diagram, modeled on a figure published in Lipmann’s 1941 article in Advances in Enzymology, shows how energy released by the metabolism of food molecules (represented by the “metabolic wheel”) is captured in the form of high-energy phosphate bonds (~P) that are used to power all other cell reactions. After the high-energy bonds are hydrolyzed, the phosphate released is recycled and reused, as indicated.
Although several molecules contain such high-energy phosphate bonds (see Panel 3–1, p. 101), it is the hydrolysis of ATP that provides the driving force for most of the energy-requiring reactions in living systems, including the contraction of muscles, the transport of substances across membranes, and the synthesis of macromolecules including proteins, nucleic acids, and carbohydrates. Indeed, in a memorial written after the death of Meyerhof in 1951, Lipmann—who would shortly win his own Nobel Prize for work on a different activated carrier—wrote: “The discovery of ATP thus was the key that opened the gates to the understanding of the conversion mechanisms of metabolic energy.”
ATP Is the Most Widely Used Activated Carrier
The most important and versatile of the activated carriers in cells is ATP (adenosine 5ʹ-triphosphate). Just as the energy stored in the raised bucket of water in Figure 3–26B can be used to drive a wide variety of hydraulic machines, ATP serves as a convenient and versatile store, or currency, of energy that can be used to drive a variety of chemical reactions in cells. As shown in Figure 3–30, ATP is synthesized in an energetically unfavorable phosphorylation reaction, in which a phosphate group is added to ADP (adenosine 5ʹ-diphosphate). When required, ATP gives up this energy packet in an energetically favorable hydrolysis to ADP and phosphate. The regenerated ADP is then available to be used for another round of the phosphorylation reaction that forms ATP, creating an ATP cycle in the cell.
More information
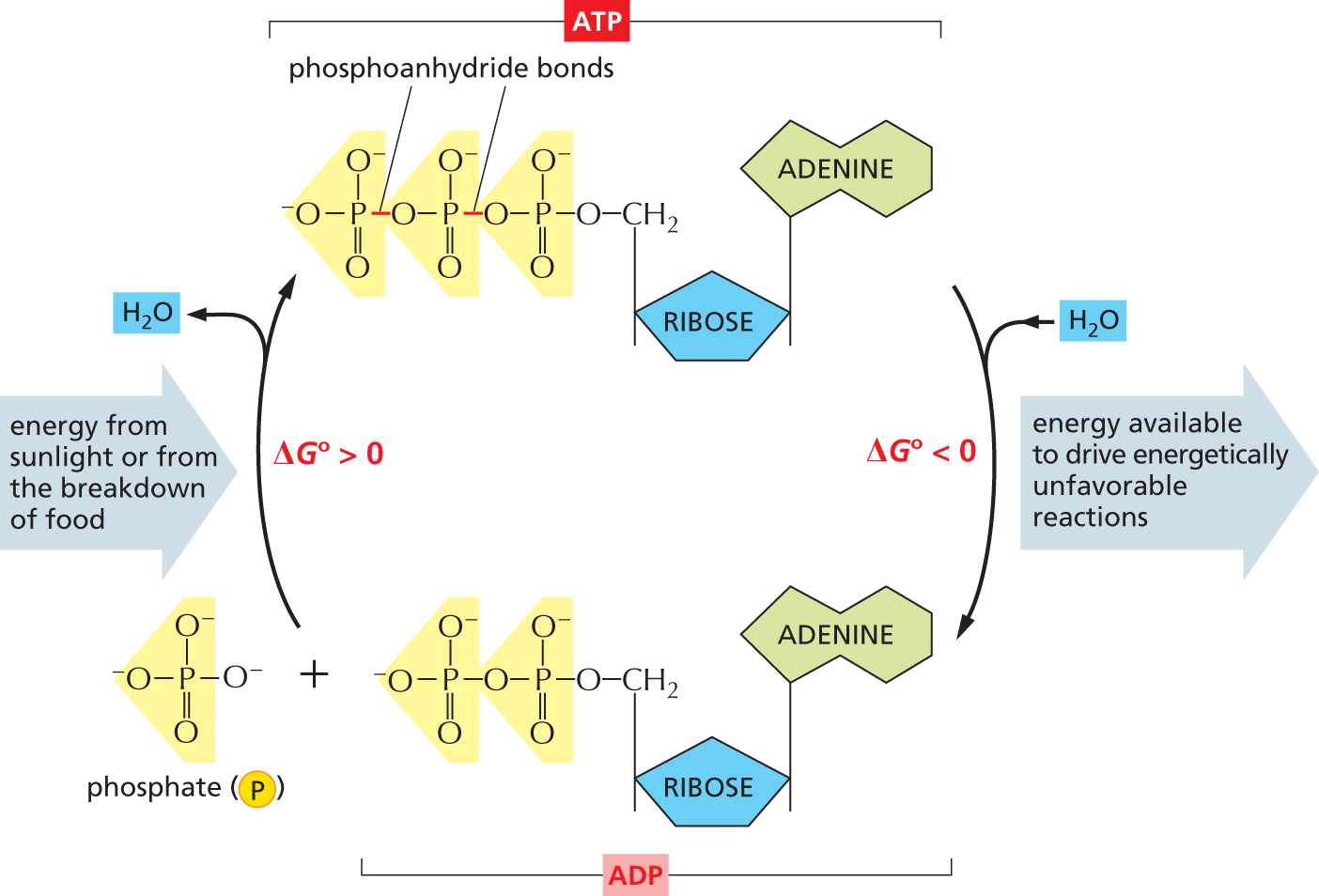
A cyclic flowchart shows the interconversion of A T P and A D P. The structure of A T P has a central ribose ring, in which C 1 is bonded to an adenine base at the top, C 4 is bonded to a methylene group at the top that is further bonded to the first phosphate group of a three phosphate group chain. The three phosphate groups are linked by phosphoanhydride bonds. A T P is converted to A D P and an inorganic phosphate molecule is released, in which the free energy, delta G 0 is less than zero and a rightward arrow pointing away from the cycle is labeled, energy available to drive energetically unfavorable reactions. The structure of A D P is similar to that of A T P, except that the methylene group is bonded to the first phosphate group of a two phosphate group chain. The structure of inorganic phosphate has a central phosphorus atom that is bonded to an oxygen anion on the left, at the top, and on the right, respectively, and it is double bonded to an oxygen atom at the bottom. A D P and the inorganic phosphate molecule combine to form A T P, in which the free energy, delta G 0 is greater than zero and a leftward arrow pointing toward the cycle is labeled, energy from sunlight or from the breakdown of food.
Figure 3–30The interconversion of ATP and ADP occurs in a cycle. The two outermost phosphate groups in ATP are held to the rest of the molecule by “high-energy” phosphoanhydride bonds and are readily transferred to other organic molecules. Water can be added to ATP to form ADP and phosphate. Inside a cell, this hydrolysis of the terminal phosphate of ATP yields between 46 and 54 kJ/mole of usable energy. (Although the ΔG° of this reaction is –30.5 kJ/mole, its ΔG inside cells is much more negative, because the ratio of ATP to the products ADP and phosphate is kept so high.)
The formation of ATP from ADP and phosphate reverses the hydrolysis reaction; because this condensation reaction is energetically unfavorable, it must be coupled to a highly energetically favorable reaction to occur.
The large negative ΔG° of the ATP hydrolysis reaction arises from a number of factors. Release of the terminal phosphate group removes an unfavorable repulsion between adjacent negative charges; in addition, the phosphate ion released is stabilized by favorable hydrogen-bond formation with water.
QUESTION 3–8
The phosphoanhydride bond that links two phosphate groups in ATP in a high-energy linkage has a ΔG° of –30.5 kJ/mole. Hydrolysis of this bond in a cell liberates from 46 to 54 kJ/mole of usable energy. How can this be? Why do you think a range of energies is given, rather than a precise number as for ΔG°?
The energetically favorable reaction of ATP hydrolysis is coupled to a large number of otherwise unfavorable reactions through which other molecules are synthesized. We will encounter several of these reactions in this chapter, where we will see exactly how this coupling is carried out. ATP hydrolysis is often accompanied by a transfer of the terminal phosphate in ATP to another molecule, as illustrated in Figure 3–31. Any reaction that involves the transfer of a phosphate group to a molecule is termed a phosphorylation reaction. A phosphorylation is an example of a condensation reaction, in which a molecule of water is released (see Figure 2–19). Phosphorylations are central to many important cell processes, including serving as key components of intracellular signaling pathways (discussed in Chapter 16).
More information
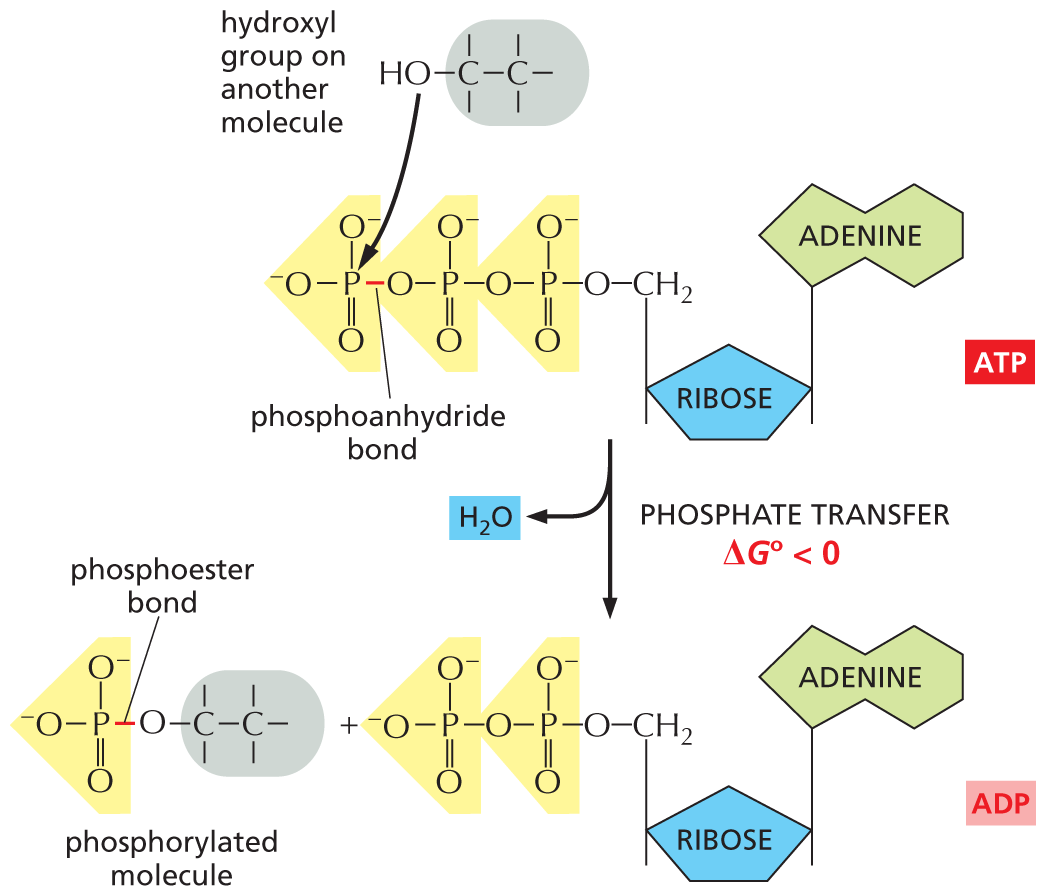
An illustration shows the transfer of a terminal phosphate group from A T P and the formation of A D P and a phosphorylated molecule. The structure of A T P has a central ribose ring, in which C 1 is bonded to an adenine base at the top, and C 4 is bonded to a methylene group at the top that is further bonded to the first phosphate group of a three phosphate group chain. The three phosphate groups are linked by phosphoanhydride bonds. A two-carbon chain molecule, in which C 1 is bonded to a hydroxyl group, attacks the phosphorus atom in the terminal phosphate group. The free energy change, delta G 0, is less than zero. The structure of A D P is similar to that of A T P, except that the methylene group is bonded to the first phosphate group of a two phosphate group chain. The structure of a phosphorylated molecule has a two-carbon chain, in which C 1 is bonded to an oxygen atom that is further bonded to the phosphorus atom of the phosphate group, and the bond between the oxygen atom and the phosphorus atom is labeled, phosphoester bond.
Figure 3–31The terminal phosphate of ATP can be readily transferred to other molecules. Because this phosphorylation reaction converts an energy-rich phosphoanhydride bond in ATP to a less energy-rich phosphoester bond in the phosphate-accepting molecule, the reaction is energetically favorable, having a large negative ΔG° (see Panel 3–1, pp. 100–101). Phosphorylation reactions of this type are involved in the synthesis of phospholipids and in the initial steps of the breakdown of sugars, as well as in many other metabolic processes.
ATP is the most abundant activated carrier in cells. It is used to supply energy for many of the pumps that actively transport substances into or out of the cell (discussed in Chapter 12) and to power the molecular motors that enable muscle cells to contract and nerve cells to transport materials along their lengthy axons (discussed in Chapter 17), to name just two important examples. Why evolution selected this particular nucleoside triphosphate over the others as the major carrier of energy, however, remains a mystery. GTP, although chemically similar to ATP, is involved in a different set of functions in the cell, as we discuss in later chapters.
Energy Stored in ATP Is Often Harnessed to Join Two Molecules Together
A common type of reaction that is needed for biosynthesis is one in which two molecules, A and B, are joined together by a covalent bond to produce A–B in an energetically unfavorable condensation reaction:
A–OH + B–H ⟶ A–B + H2O
ATP hydrolysis can be coupled indirectly to this reaction to make it go forward. In this case, energy from ATP hydrolysis is first used to convert A–OH to a higher-energy intermediate compound, which then reacts directly with B–H to give A–B. The simplest mechanism involves the transfer of a phosphate from ATP to A–OH to make A–O–PO3, in which case the reaction pathway contains only two steps (Figure 3–32A). The condensation reaction, which by itself is energetically unfavorable, has been forced to occur by being coupled to ATP hydrolysis in an enzyme-catalyzed reaction pathway.
More information
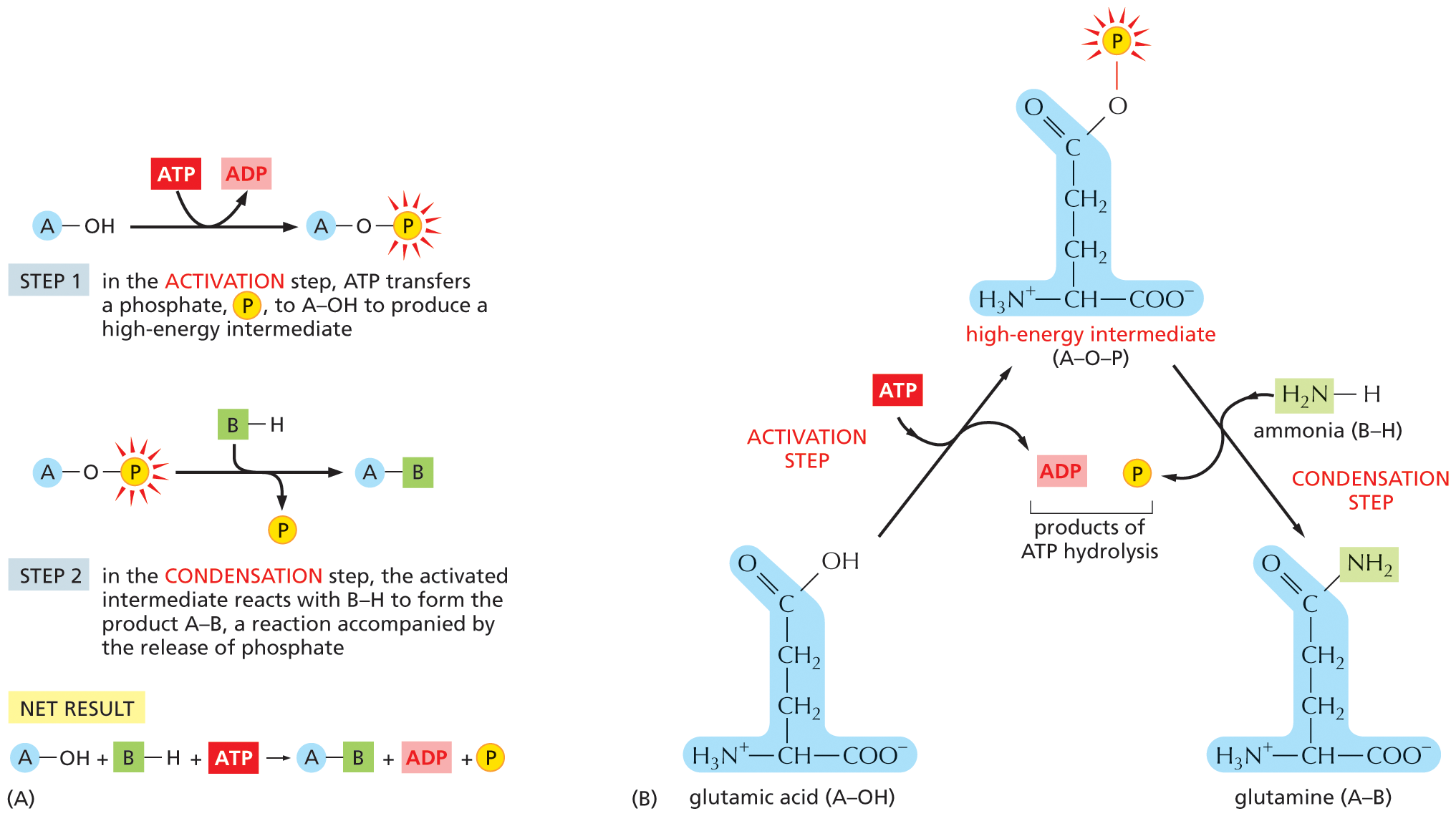
A two-part illustration, A and B, shows a schematic of a biosynthetic reaction and a specific example, respectively. Section A: Step 1: A molecule A is bonded to a hydroxyl group, which undergoes a reaction during which A T P is converted to A D P and an intermediate is formed, in which the molecule A is bonded to an oxygen atom that is further bonded to a phosphate group.
Step 2: The intermediate undergoes a reaction in the presence of B H to form a product A B and inorganic phosphate is released during the reaction.
Net Result: A molecule A is bonded to a hydroxyl group and a molecule B is bonded to a hydrogen atom, and an A T P molecule undergoes a reaction to form a product A B, A D P, and inorganic phosphate.
Section B: Glutamic acid, represented by a molecule A bonded to a hydroxyl group, undergoes an activation step, during which A T P is consumed and A D P is released. The high-energy intermediate, A O P formed has a structure similar to that of the glutamic acid, except that the oxygen atom in the hydroxyl group is bonded to a phosphate group. This high-energy intermediate undergoes a condensation step, in which it reacts with a molecule of ammonia, represented by a molecule B bonded to a hydrogen atom; and a molecule of inorganic phosphate is released. Glutamine, the product formed, has a similar structure to that of glutamic acid, except that the carbon atom of carboxylic acid is double bonded to an oxygen atom and to an amino group.
Figure 3–32How an energetically unfavorable biosynthetic reaction can be driven by ATP hydrolysis. (A) Schematic illustration of the reactions described in the text. In this set of reactions, a phosphate group is first donated by ATP to form a high-energy intermediate, A–O–PO3, which then reacts with the other substrate, B–H, to form the product A–B. (B) The biosynthesis of the amino acid glutamine from glutamic acid. Glutamic acid, which corresponds to the A–OH shown in (A), is first converted to a high-energy phosphorylated intermediate, which corresponds to A–O–PO3. This intermediate then reacts with ammonia (which corresponds to B–H) to form glutamine. In this example, both steps occur on the surface of the same enzyme, glutamine synthetase (not shown). ATP hydrolysis can drive this energetically unfavorable reaction because it produces a favorable free-energy change (ΔG° of –30.5 kJ/mole) that is larger in magnitude than the energy required for the synthesis of glutamine from glutamic acid plus NH3 (ΔG° of +14.2 kJ/mole). For clarity, the glutamic acid side chain is shown in its uncharged form.
A biosynthetic reaction of exactly this type is employed to synthesize the amino acid glutamine, as illustrated in Figure 3–32B. We will see later in this chapter that very similar (but more complex) mechanisms are also used to produce nearly all of the large molecules of the cell.
NADH and NADPH Are Both Activated Carriers of Electrons
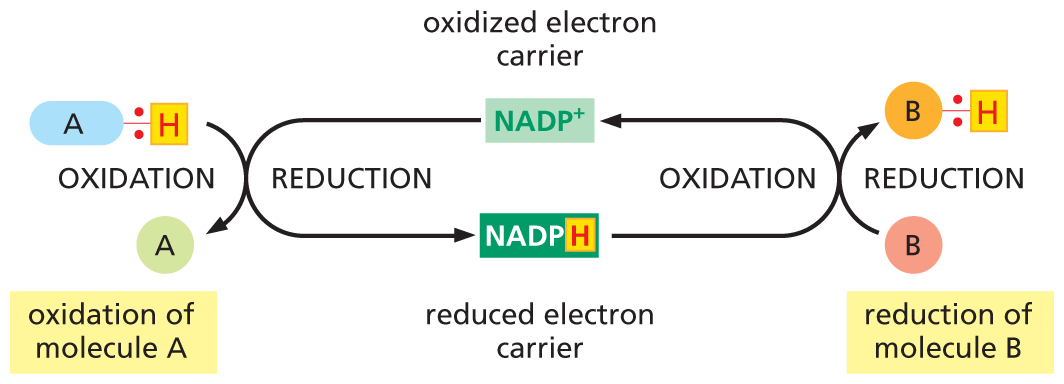
While ATP carries phosphate groups in a transferrable, “high-energy” linkage, other activated carriers are specialized to carry “high-energy” electrons and hydrogen atoms. The most important of these electron carriers are NADH (nicotinamide adenine dinucleotide) and the closely related molecule NADPH (nicotinamide adenine dinucleotide phosphate). Both NADH and NADPH carry energy in the form of two high-energy electrons plus a proton (H+), which together form a hydride ion (H–). When these activated carriers pass their hydride ion to a donor molecule, they become oxidized to form NAD+ and NADP+, respectively.
Like ATP, NADPH is an activated carrier that participates in many important biosynthetic reactions that would otherwise be energetically unfavorable. NADPH is produced according to the general scheme shown in Figure 3–33. During a special set of energy-yielding catabolic reactions, a hydride ion is removed from the substrate molecule and added to the nicotinamide ring of NADP+ to form NADPH. This is a typical oxidation–reduction reaction: the substrate is oxidized and NADP+ is reduced.
More information
A flow diagram illustrates coupled oxidation and reduction. N A D P plus, labeled oxidized electron carrier, undergoes reduction to form N A D P H, during which a molecule A bonded to a hydrogen atom undergoes oxidation to form molecule A and the hydrogen atom is bonded to N A D P plus and forms N A D P H. N A D P H, labeled reduced electron carrier, undergoes oxidation, to form N A D P plus. During this reaction, a molecule B undergoes reduction and forms a molecule B bonded to a hydrogen atom.
Figure 3–33NADPH is an activated carrier of electrons that participates in oxidation–reduction reactions. NADPH is produced in reactions of the general type shown on the left, in which two electrons are removed from a substrate (A–H). The oxidized form of the carrier molecule, NADP+, receives these two electrons as one hydrogen atom plus an electron (a hydride ion). Because NADPH holds its hydride ion in a high-energy linkage, this ion can easily be transferred to other molecules, such as B, as shown on the right. In this reaction, NADPH is reoxidized to yield NADP+, thus completing the cycle.
The hydride ion carried by NADPH is given up readily in a subsequent oxidation–reduction reaction, because the nicotinamide ring can achieve a more stable arrangement of electrons without it (Figure 3–34). In this subsequent reaction, which regenerates NADP+, the NADPH becomes oxidized and the substrate becomes reduced—thus completing the NADPH cycle (see Figure 3–33). NADPH is efficient at donating its hydride ion to other molecules for the same reason that ATP readily transfers a phosphate: in both cases, the transfer is accompanied by a large negative free-energy change. One example of the use of NADPH in biosynthesis is shown in Figure 3–35.
More information
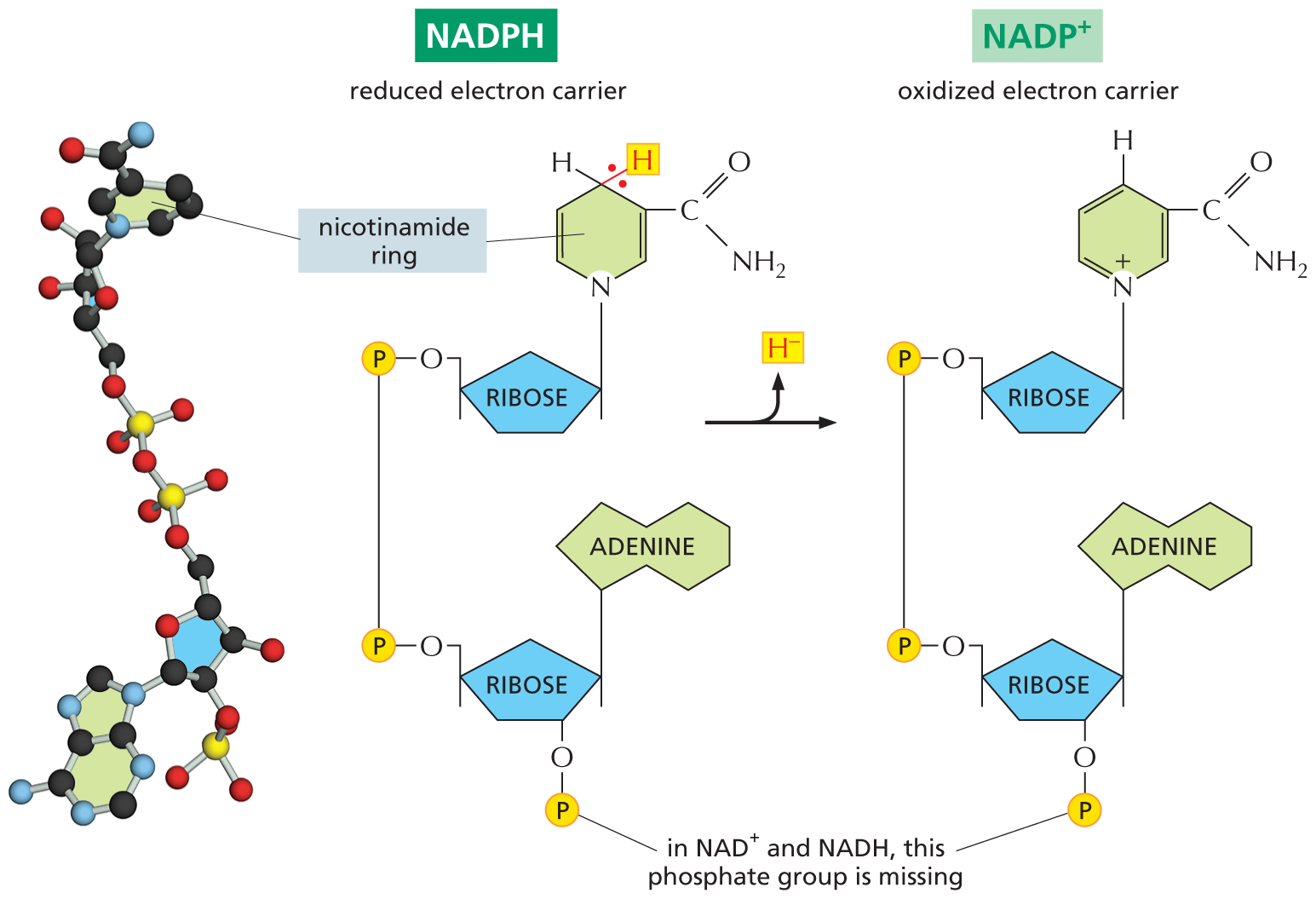
A ball and stick model of N A D P H next to a schematic N A D P H undergoing a reaction to produce N A D H. The structure of N A D P H has two ribose rings arranged vertically. The ribose at the top has the following substituents: C 1 is bonded to a nicotinamide ring at the top, and C 4 is bonded to an oxygen atom that is further bonded to a phosphate group. This phosphate group is bonded to a phosphate group at the bottom that is bonded to an oxygen atom. The nicotinamide ring has a cyclohexane ring, in which the first position is occupied by a nitrogen atom, C 2 is double bonded to C 3, and C 5 is double bonded to C 6. In the ring, C 3 is bonded to a carbonyl group and the carbonyl carbon is further bonded to an amino group, and C 4 is bonded to two hydrogen atoms, of which one of them is highlighted and the two electrons involved in the single bond are depicted. The ribose ring at the bottom has the following substituents: C 1 is bonded to adenine ring at the top, C 2 is single bonded to an oxygen atom at the bottom that is further bonded to a phosphate group, represented by a yellow sphere labeled P, and C 4 is bonded to an oxygen atom that is further bonded to a phosphate group, which links the first and second ribose ring. N A D P H loses the highlighted hydrogen atom and forms N A D P plus, which has a similar structure to that of N A D P H, except that the nitrogen atom in the nicotinamide ring carries a positive charge. A text pointing toward the phosphate group bonded to C 3 of the second ribose ring reads, in N A D plus and N A D H, this phosphate group is missing.
Figure 3–34NADPH accepts and donates electrons via its nicotinamide ring. NADPH donates its “high-energy” electrons together with a proton (the equivalent of a hydride ion, H–). This reaction, which oxidizes NADPH to NADP+, is energetically favorable because the nicotinamide ring is more stable when these electrons are absent. The ball-and-stick model on the left shows the structure of NADP+. NAD+ and NADH are identical in structure to NADP+ and NADPH, respectively, except that they lack the phosphate group, as indicated.
More information
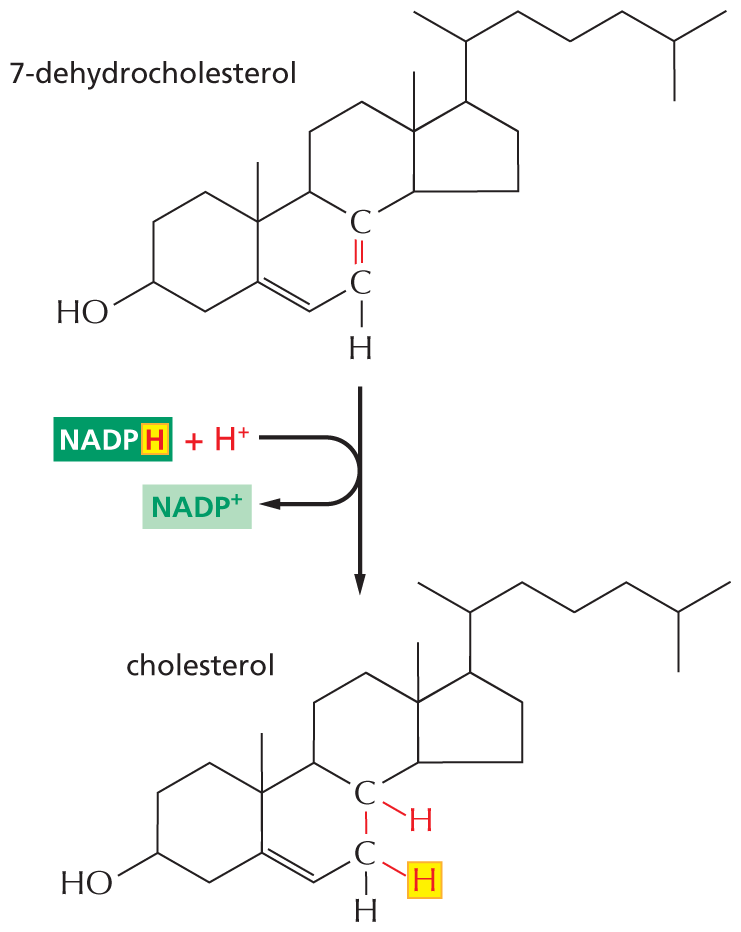
An illustration shows the reduction of 7 dehydrocholesterol to form cholesterol. The skeletal line structure of 7-dehydrocholesterol has four fused ring structures of which the first, second, and third rings are six membered, and the fourth ring is five membered. The carbon atoms are numbered according to IUPAC nomenclature. C 3 is bonded to a hydroxyl group. C 5 is double bonded to C 6. C 7 is double bonded to C 8, which is highlighted and C 7 is bonded to a hydrogen atom. C 10 is bonded to a methyl group. C 13 is bonded to a methyl group. C 17 is bonded to C 2 of a seven-carbon chain, in which C 6 is bonded to a methyl group. 7-dehydrocholesterol undergoes a reduction reaction, during which N A D P H, with highlighted H and a highlighted proton enters the reaction and N A D P plus is released, and cholesterol is formed. The skeletal line structure of cholesterol is similar to that of 7-dehydrocholesterol, except that the highlighted double bond is converted to a single bond, C 8 is bonded to a hydrogen atom that entered the reaction as a proton, and C 7 is bonded to highlighted hydrogen from N A D P H.
Figure 3–35NADPH participates in the final stage of one of the biosynthetic routes leading to cholesterol. As in many other biosynthetic reactions, the reduction of the C=C bond is achieved by the transfer of a hydride ion from the activated carrier NADPH, plus a proton (H+) from solution.
NADPH and NADH Have Different Roles in Cells
NADPH and NADH differ in a single phosphate group, which is located far from the region involved in electron transfer in NADPH (see Figure 3–34). Although this phosphate group has no effect on the electron-transfer properties of NADPH compared with NADH, it is nonetheless crucial for their distinctive roles, as it gives NADPH a slightly different shape from NADH. This subtle difference in conformation makes it possible for the two carriers to bind as substrates to different sets of enzymes and thereby deliver electrons (in the form of hydride ions) to different target molecules.
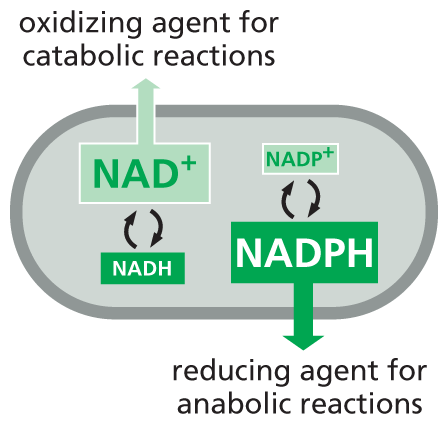
Why should there be this division of labor? The answer lies in the need to regulate two sets of electron-transfer reactions independently. NADPH operates chiefly with enzymes that catalyze anabolic reactions, supplying the high-energy electrons needed to synthesize energy-rich biological molecules. NADH, by contrast, has a special role as an intermediate in the catabolic system of reactions that generate ATP through the oxidation of food molecules, as we discuss in Chapter 13. The genesis of NADH from NAD+ and that of NADPH from NADP+ occurs by different pathways that are independently regulated, so that the cell can adjust the supply of electrons for these two contrasting purposes. Inside the cell, the ratio of NAD+ to NADH is kept high, whereas the ratio of NADP+ to NADPH is kept low. This arrangement provides plenty of NAD+ to act as an oxidizing agent and plenty of NADPH to act as a reducing agent—as required for their special roles in catabolism and anabolism, respectively (Figure 3–36).
More information
An illustration shows the role of N A D plus and N A D P H in a cell. The illustration shows the outline of a cell as an oval structure. N A D and N A D H are interconverted and N A D plus is an oxidizing agent for catabolic reactions. N A D P H and N A D P plus are interconverted and N A D P H is a reducing agent for anabolic reactions.
Figure 3–36NADPH and NADH have different roles in the cell, and the relative concentrations of these carrier molecules influence their affinity for electrons. Keeping reduced NADPH at a higher concentration than its oxidized counterpart, NADP+, makes NADPH a stronger electron donor. This arrangement ensures that NADPH can serve as a reducing agent for anabolic reactions. The reverse is true for NADH. Cells keep the amount of reduced NADH lower than that of NAD+, which makes NAD+ a better electron acceptor. Thus NAD+ acts as an effective oxidizing agent, accepting electrons generated during the oxidative breakdown of food molecules, as discussed in Chapter 13.
Cells Make Use of Many Other Activated Carriers
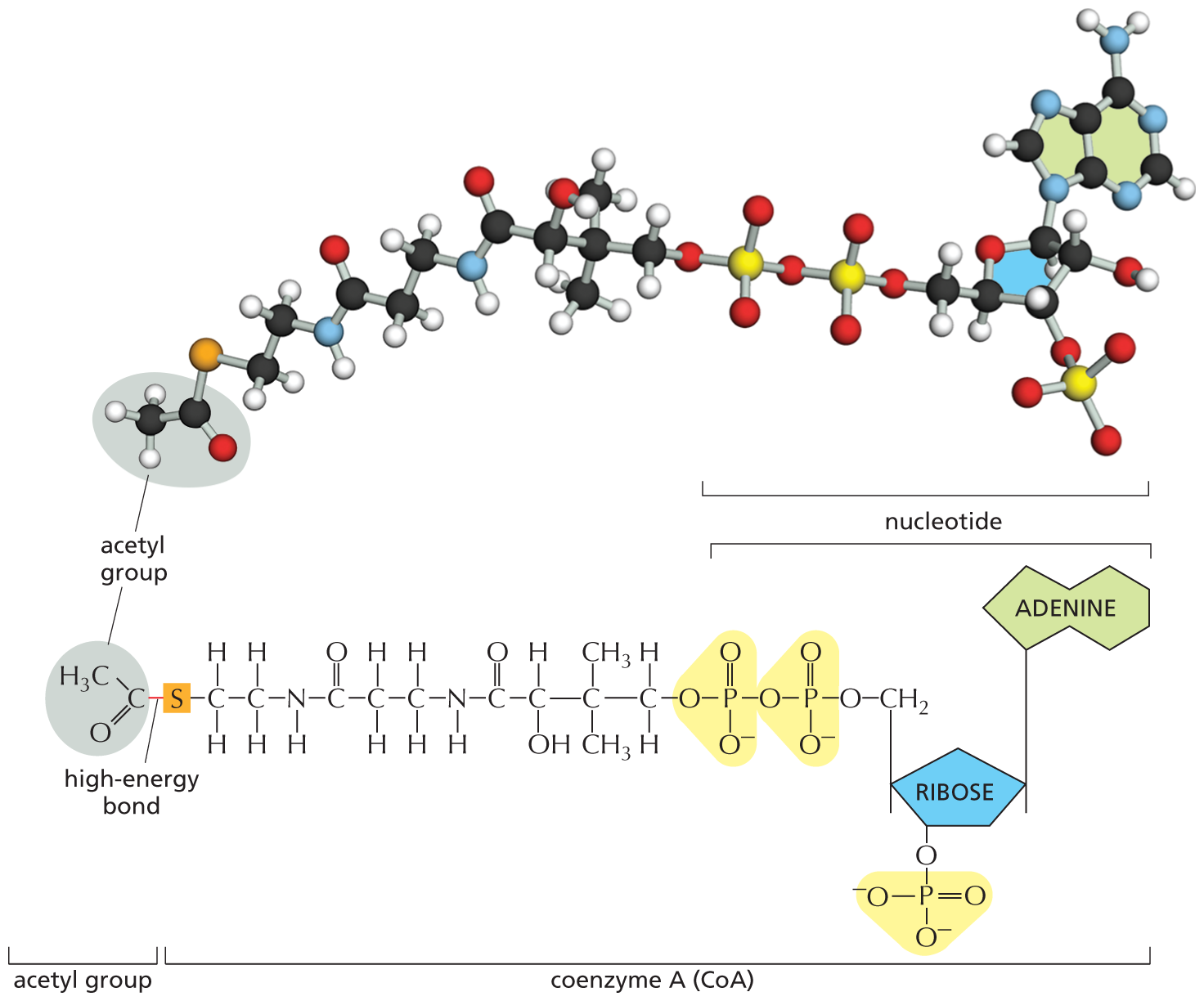
In addition to ATP (which transfers a phosphate) and NADPH and NADH (which transfer electrons and hydrogen), cells make use of other activated carriers that pick up and carry a chemical group in an easily transferred, high-energy linkage. FADH2, like NADH and NADPH, carries hydrogen and high-energy electrons (see Figure 13–13B). But other important reactions involve the transfers of acetyl, methyl, carboxyl, and glucose groups from activated carriers for the purpose of biosynthesis (Table 3–2). Coenzyme A, for example, can carry an acetyl group in a readily transferable linkage. This activated carrier, called acetyl CoA (acetyl coenzyme A), is shown in Figure 3–37. It is used, for example, to sequentially add two-carbon units in the biosynthesis of the hydrocarbon tails of fatty acids.
TABLE 3–2 SOME ACTIVATED CARRIERS WIDELY USED IN METABOLISM
Activated Carrier
Group Carried in “High-Energy” Linkage
ATP
Phosphate
NADH, NADPH, FADH2
Electrons and hydrogens
Acetyl CoA
Acetyl group
Carboxylated biotin
Carboxyl group
S-Adenosylmethionine
Methyl group
Uridine diphosphate glucose
Glucose
More information
An illustration shows a ball and stick model and structural formula of acetyl coenzyme A. The structural formula of acetyl coenzyme A has a central ribose ring, in which C 1 is bonded to an adenine ring at the top, C 3 is bonded to one of the oxygen atoms in a highlighted phosphate group, and C 4 is bonded to methylene that is bonded to highlighted diphosphate anion, which is further bonded to an oxygen atom that is bonded to a 11-atom chain in the following order: C 1 is a methylene group, C 2 is bonded to two methyl groups at the top and at the bottom, respectively, C 3 is bonded to a hydrogen atom at the top and to a hydroxyl group at the bottom, C 4 is a carbonyl group, C 5 is replaced by an N H group, C 6 and C 7 are each methylene groups, C 8 is a carbonyl group, C 9 is replaced by an N H group, and C 10 and C 11 are each methylene groups. C 11 is bonded to a sulfur atom on its left that is further single bonded to a carbonyl group and the bond is labeled high-energy bond, and the carbonyl carbon is bonded to a methyl group. In the structural formula, the ribose ring, the adenine base, and the highlighted diphosphate anion are labeled together as nucleotide; the ribose ring, the adenine base, the highlighted diphosphate anion, and the 11-carbon chain are labeled together as coenzyme A (Co A), and the terminal carbonyl group bonded to a methyl group is labeled acetyl group. The ball and stick model of acetyl coenzyme A is shown at the top.
Figure 3–37Acetyl coenzyme A (CoA) is another important activated carrier. A ball-and-stick model is shown above the structure of acetyl CoA. The sulfur atom (orange) forms a thioester bond to acetate. Because the thioester bond releases a large amount of free energy when it is hydrolyzed, it is said to be a "high-energy" linkage. Thus the acetyl group carried by CoA can be readily transferred to other molecules.
For acetyl CoA and the other activated carriers in Table 3–2, the transferable group makes up only a small part of the molecule. The rest consists of a large organic portion that serves as a convenient “handle,” facilitating the recognition of the carrier molecule by specific enzymes. As with acetyl CoA, this handle portion very often contains a nucleotide. This curious fact may be a relic from an early stage of cell evolution. It is thought that the main catalysts for early life-forms on Earth were RNA molecules (or their close relatives) and that proteins were a later evolutionary addition. It is therefore tempting to speculate that many of the activated carriers that we find today originated in an earlier RNA world, where their nucleotide portions would have been useful for binding these carriers to RNA-based catalysts, or ribozymes (discussed in Chapter 7).
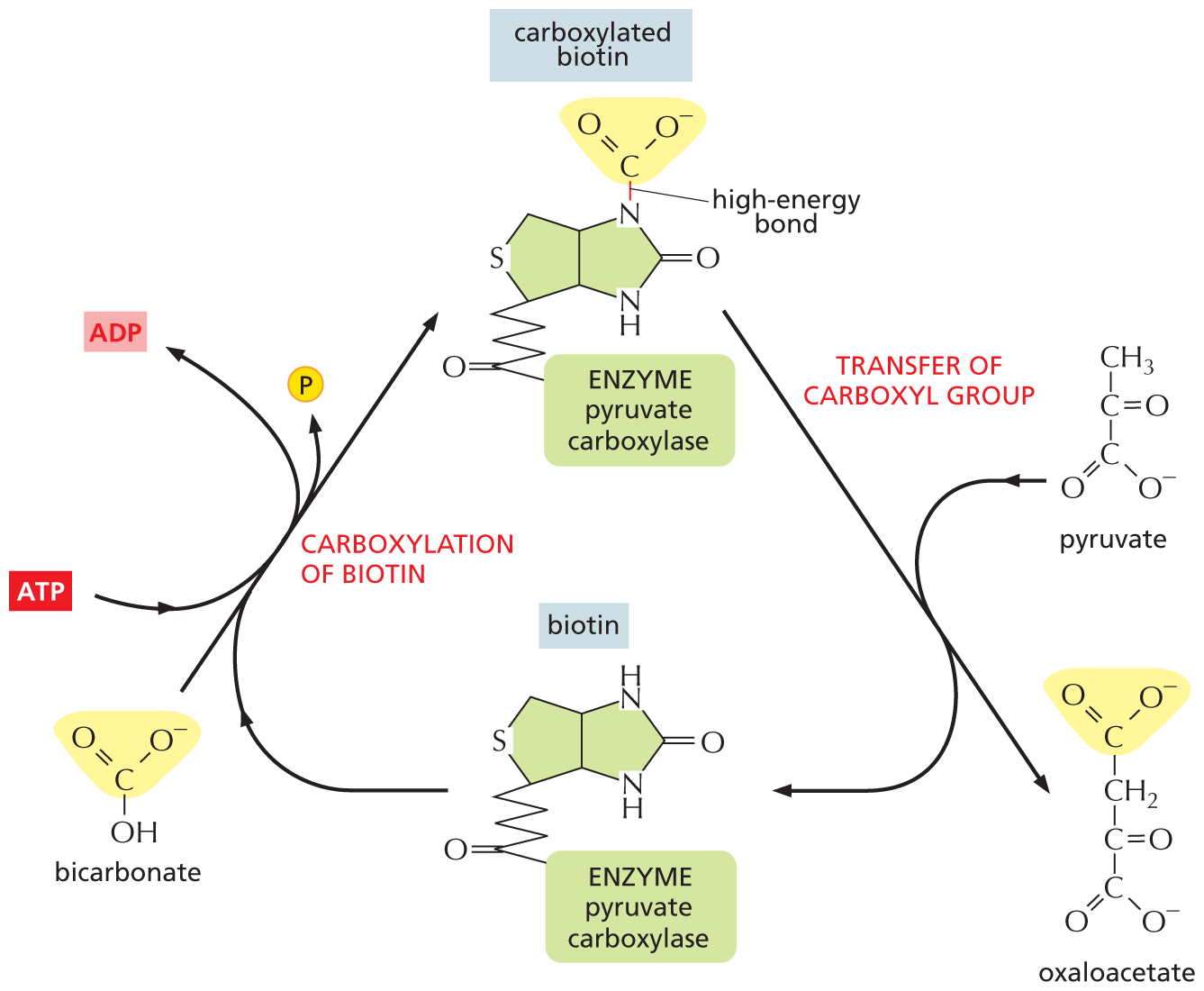
Activated carriers are usually generated in reactions coupled to ATP hydrolysis, as shown for biotin in Figure 3–38. Therefore, the energy that enables their groups to be used for biosynthesis ultimately comes from the catabolic reactions that generate ATP. This also applies to the synthesis of large macromolecules—nucleic acids, proteins, and polysaccharides—as we discuss next.
More information
An illustration depicts the reaction mechanism for the transfer of a carboxyl group from bicarbonate to pyruvate by the action of biotin. The structure of biotin has two cyclopentane rings fused horizontally at the fourth and fifth carbon atoms. In the first ring on the left, the first position is replaced by a sulfur atom, C 2 of the ring is bonded to C 1 of a five-carbon skeletal chain, in which C 5 is a carbonyl group and it is further bonded to enzyme pyruvate carboxylase. Bicarbonate has a central carbon atom that is single bonded to an oxygen anion on the right, a hydroxyl group at the bottom, and double bonded to an oxygen atom on the left. Bicarbonate undergoes carboxylation of biotin, in which biotin enters the reaction and a phosphate group, represented by a yellow sphere labeled P, is released. During the reaction, A T P is converted to A D P. A carboxylated biotin is formed, which has a structure similar to biotin, except that N 1 is single bonded to the central carbon of bicarbonate, which is labeled high-energy bond, and the carbon atom is bonded to an oxygen anion and double bonded to oxygen atom. The carboxylate biotin undergoes transfer of carboxyl group and forms oxaloacetate, and during the reaction pyruvate enters the reaction and biotin is formed again. Oxaloacetate has a four-carbon vertical chain, in which C 1 and C 4 are each a carboxylate anion and C 1 is highlighted, C 2 is a methylene group, and C 3 is a carbonyl group.
Figure 3–38Biotin transfers a carboxyl group to a substrate. A number of enzymes use biotin to transfer a carboxyl group to a substrate. Shown here is the reaction in which biotin, held by the enzyme pyruvate carboxylase, accepts a carboxyl group from bicarbonate and transfers it to pyruvate, producing oxaloacetate, a molecule required in the citric acid cycle (discussed in Chapter 13). Other enzymes use biotin to transfer carboxyl groups to other molecules. Note that the synthesis of carboxylated biotin requires energy derived from ATP hydrolysis—a general feature that applies to many activated carriers. Because biotin must be derived from the diet, it is considered a vitamin. This is true for several of the activated carriers used by cells.
The Synthesis of Biological Polymers Requires an Energy Input
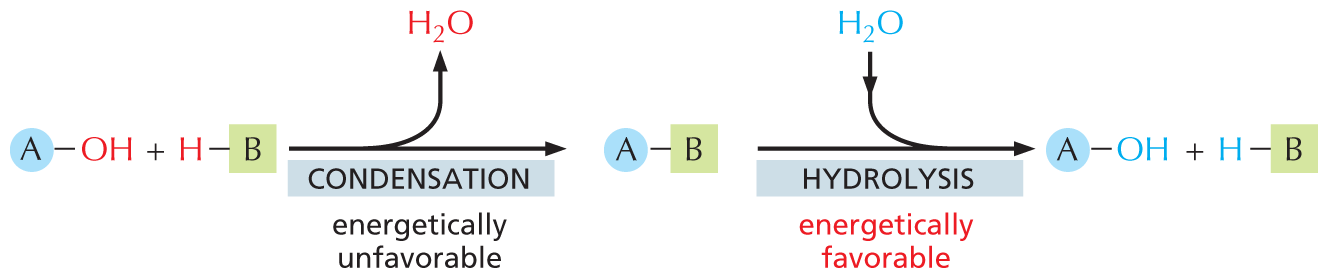
The macromolecules of the cell constitute the vast majority of its dry mass—that is, the mass not due to water. These molecules are made from subunits (or monomers) that are linked together by bonds formed during an enzyme-catalyzed condensation reaction. The reverse reaction—the breakdown of polymers—occurs through enzyme-catalyzed hydrolysis reactions. These hydrolysis reactions are energetically favorable, whereas the corresponding biosynthetic reactions require an energy input and are more complex (Figure 3–39).
More information
An illustration shows macromolecular synthesis and degradation. In step 1, a molecule A bonded to a hydroxyl group reacts with a molecule B bonded to a hydrogen atom and undergoes an energetically unfavorable condensation reaction, to form a product where molecule A is bonded to B. A water molecule is released during the reaction. In step 2, the molecule A bonded to B undergoes an energetically favorable hydrolysis, during which a water molecule enters the reaction and the initial reactants are formed.
Figure 3–39In cells, macromolecules are synthesized by condensation reactions and broken down by hydrolysis reactions. Condensation reactions are all energetically unfavorable, whereas hydrolysis reactions are all energetically favorable.
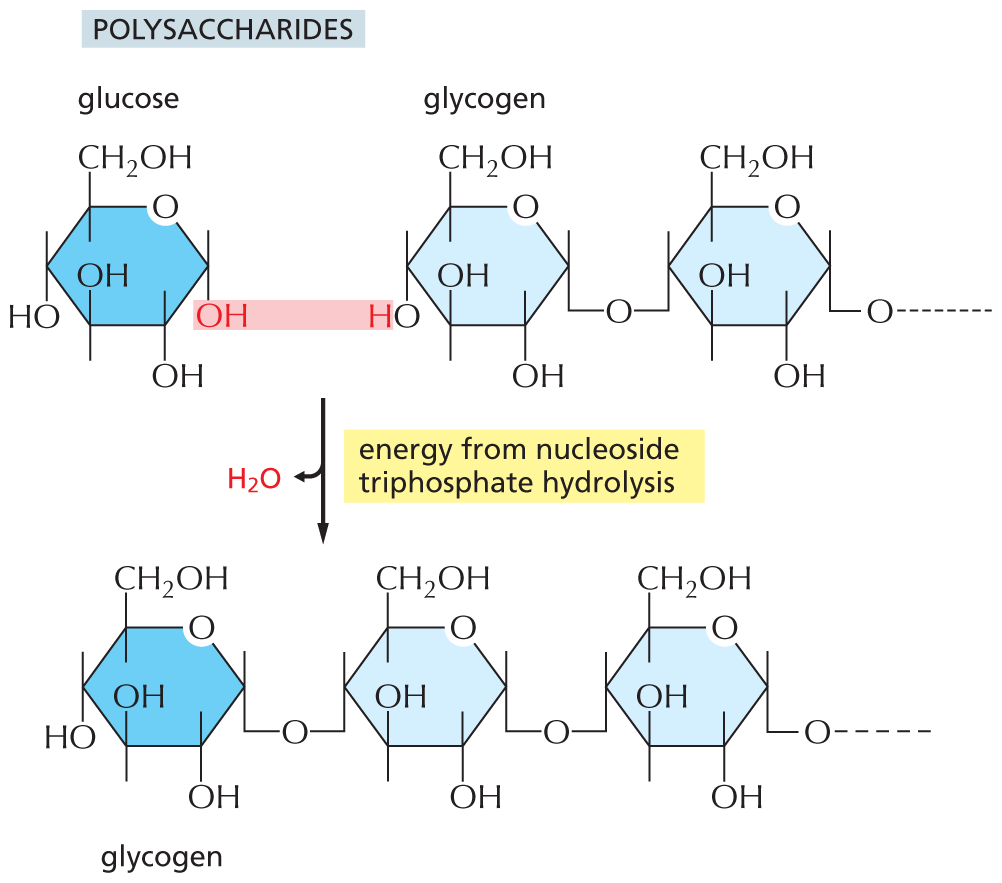
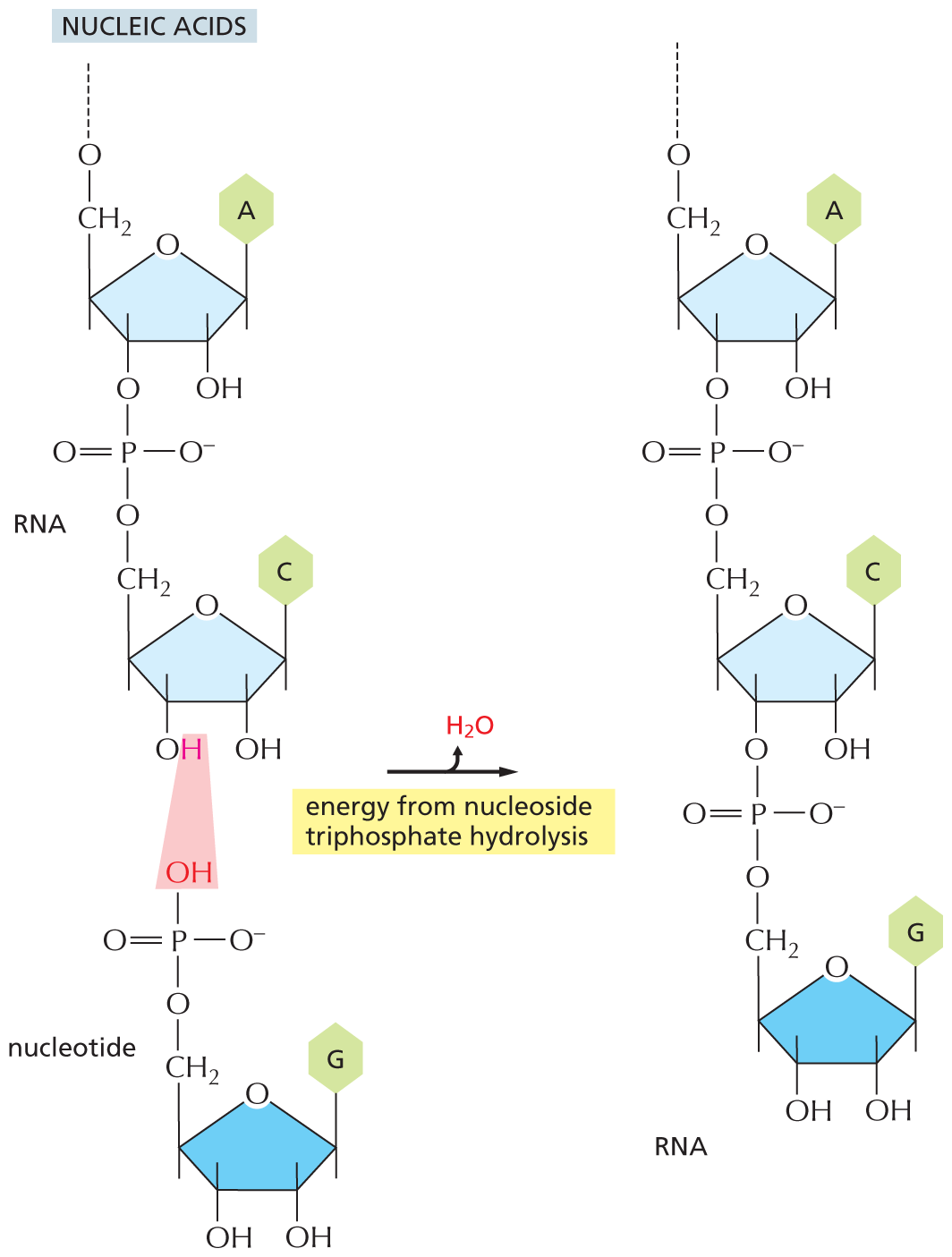
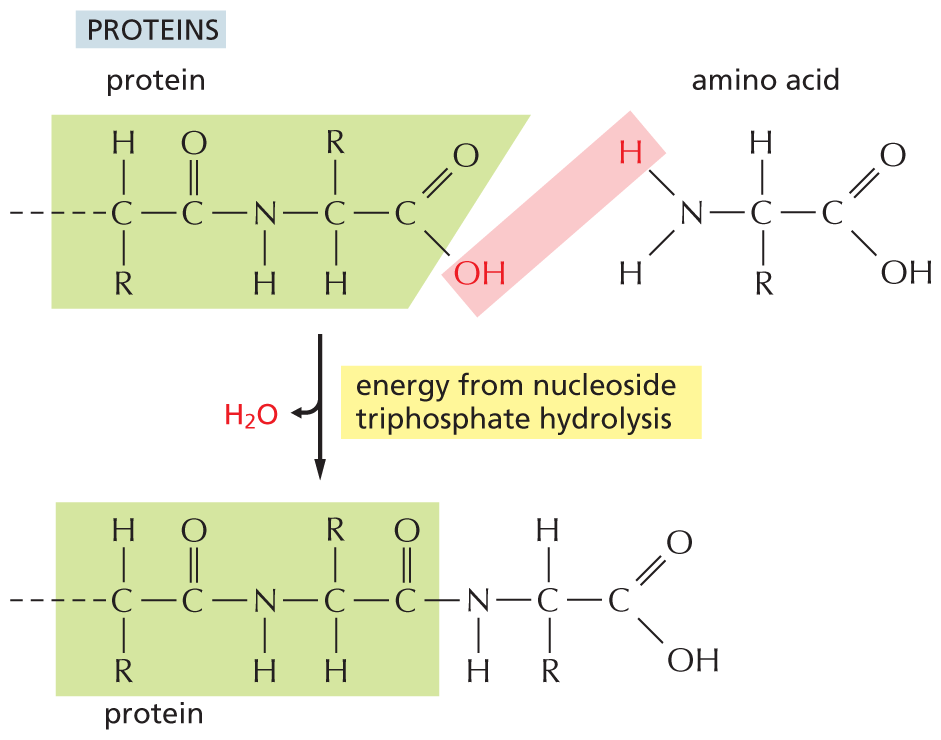
The nucleic acids (DNA and RNA), proteins, and polysaccharides are all polymers that are produced by the repeated addition of a subunit onto one end of a growing chain. The mode of synthesis of each of these macromolecules is outlined in Figure 3–40. As indicated, the condensation step in each case depends on energy provided by the hydrolysis of a nucleoside triphosphate. And yet, except for the nucleic acids, there are no phosphate groups left in the final product molecules. How, then, is the energy of ATP hydrolysis coupled to polymer synthesis?
More information
An illustration shows the synthesis of polysaccharides from glucose and glycogen, via release of water and energy input from nucleoside triphosphate hydrolysis. Glucose and glycogen undergoes a condensation reaction to add a glucose molecule to glycogen chain. A water molecule is released during the reaction. The reaction is mediated by the energy from nucleoside triphosphate hydrolysis. The structure of glucose has a pyranose ring, in which the first position is occupied by an oxygen atom, C 1, C 2, and C 4 are each bonded to a hydroxyl group at the bottom, C 3 is bonded to a hydroxyl group at the top, and C 5 is bonded to a hydroxymethyl group at the top. The hydroxyl group bonded to C 1 is highlighted. In the structure of glycogen, two terminal ring structures are shown. Both the rings have a central pyranose ring and have similar structures. In the pyranose ring, the first position is occupied by an oxygen atom, C 1 of the first ring is bonded to an oxygen atom that is further bonded to C 4 of the second ring and links the two ring structures and C 1 of the second ring is bonded to an oxygen atom that is further bonded to the remaining chain of glycogen. C 2 is bonded to a hydroxyl group at the bottom and C 3 is bonded to a hydroxyl group at the top. C 4 of the first ring is bonded to a hydroxyl group at the bottom, in which the hydrogen atom is highlighted and C 4 of the second ring is bonded to an oxygen atom at the bottom, which links the first and second ring. C 5 of both the rings is bonded to a hydroxymethyl group at the top. The highlighted hydroxyl group in glucose and highlighted hydrogen atom in glycogen are removed as water. The product formed, glycogen has three terminal ring structures and the structure of the new ring is identical to the previous two rings.
(A)More information
An illustration shows the synthesis of nucleic acids from an R N A chain and a nucleotide, via release of water and energy input from nucleoside triphosphate hydrolysis. A nucleotide and an R N A chain undergoes a condensation reaction to add a nucleotide molecule to the R N A chain. A water molecule is released during the reaction. The reaction is mediated by the energy from nucleoside triphosphate hydrolysis. The structure of R N A chain and nucleotide are shown vertically. In the structure of R N A, two terminal rings are shown. Both the rings have a central furanose ring and have similar structures. In the furanose ring, first position is occupied by an oxygen atom, C 1 of the first ring is bonded to adenine base at the top and C 1 of the second ring is bonded to cytosine base at the top. C 2 of both the rings is bonded to a hydroxyl group at the bottom. C 3 of the first ring is bonded to an oxygen atom of a phosphate group, which links the first and the second rings. C 3 of the second ring is bonded to a hydroxyl group, in which the hydrogen atom is highlighted. C 4 is bonded to a methylene group at the top that is further bonded to an oxygen atom, which is further bonded to the remaining chain of R N A. C 4 of the second ring is bonded to a methylene group at the top that is further bonded to an oxygen atom in the phosphate group linking the first and the second ring. In the phosphate group, the central phosphorus atom is single bonded to an oxygen atom at the top bonded to C 3 of the first ring, single bonded to an oxygen anion on the right, single bonded to an oxygen atom at the bottom that is bonded to methylene group of second ring, and double bonded to an oxygen atom on the left. The structure of the nucleotide has a pyranose ring, in which C 1 is bonded to guanine base at the top, C 2 and C 3 are each bonded to a hydroxyl group at the bottom, C 4 is bonded to a methylene group at the top, which is single bonded to an oxygen atom of phosphate group. In the phosphate group, the central phosphorus atom is single bonded to a highlighted hydroxyl group at the top, single bonded to an oxygen anion on the right, single bonded to an oxygen atom at the bottom that is bonded to methylene group of the second ring, and double bonded to an oxygen atom on the left. The highlighted hydrogen atom in the first ring and the highlighted hydroxyl group in the second ring are removed as water molecule. The product formed, an R N A chain with an additional nucleotide group has a similar structure as the reactant R N A chain.
(B)More information
An illustration shows the synthesis of proteins from a protein chain and an amino acid, via release of water and energy input from nucleoside triphosphate hydrolysis. An amino acid and a protein chain undergoes a condensation reaction to add an amino acid molecule to the protein chain. A water molecule is released during the reaction. The reaction is mediated by the energy from nucleoside triphosphate hydrolysis. The structure of the protein chain from left is as follows: a carbon atom is bonded to a hydrogen atom at the top, a carbonyl group on the right, an R group at the bottom, and to the rest of the protein chain on the left. The carbonyl group is bonded to an N H group on the right that is further bonded to a carbon atom. The carbon atom is bonded to an R group at the top, a carboxylic acid group on the right, and to a hydrogen atom at the bottom. In the carboxylic acid group, the O H group is highlighted. The general formula of an amino acid is shown and it has a central carbon atom that is bonded to a hydrogen atom at the top, a carboxylic acid group on the right, an R group at the bottom, and to an amino group on the left. In the amino group, one of the hydrogen atoms is highlighted, and the highlighted hydroxyl group in the protein chain and the highlighted hydrogen atom are removed as water. The protein chain is linked to the amino acid.
(C)Figure 3–40The synthesis of macromolecules requires an input of energy. Synthesis of a portion of (A) a polysaccharide, (B) a nucleic acid, and (C) a protein is shown here. In each case, synthesis involves a condensation reaction in which water is lost; the atoms involved are shaded in pink. Not shown is the consumption of high-energy nucleoside triphosphates that is required to activate each subunit prior to its addition. In contrast, the reverse reaction—the breakdown of all three types of polymers—occurs through the simple addition of water, or hydrolysis (not shown).
Each type of macromolecule is generated by an enzyme-catalyzed pathway that resembles the one discussed previously for the synthesis of the amino acid glutamine (see Figure 3–32). The principle is exactly the same, in that the –OH group that will be removed in the condensation reaction is first activated by forming a high-energy linkage to a second molecule. The mechanisms used to link ATP hydrolysis to the synthesis of proteins and polysaccharides, however, are more complex than that used for glutamine synthesis. In the biosynthetic pathways leading to these macromolecules, several high-energy intermediates are consumed in series to generate the final high-energy bond that will be broken during the condensation step. One important example of such a biosynthetic reaction, that of protein synthesis, is discussed in detail in Chapter 7.
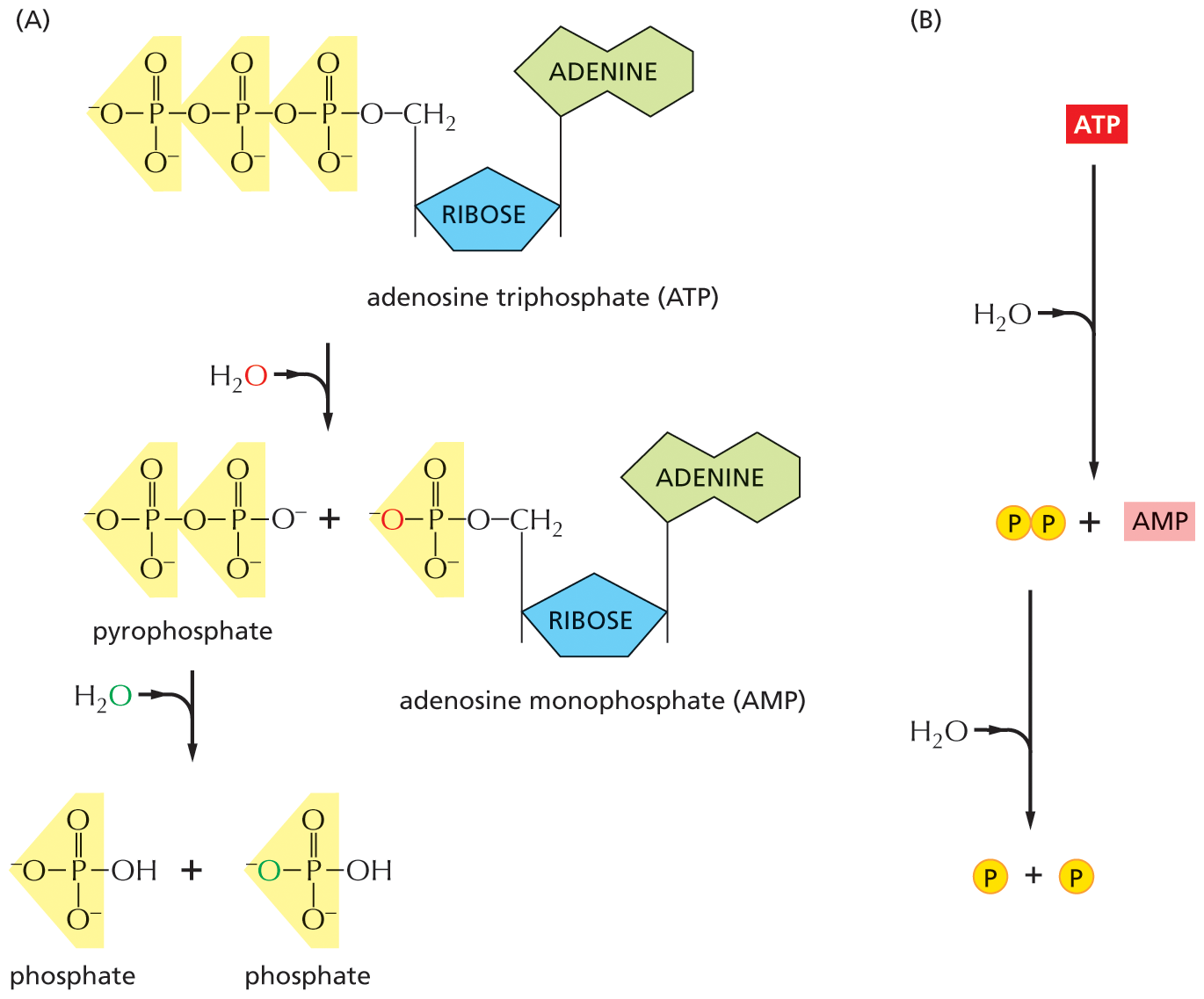
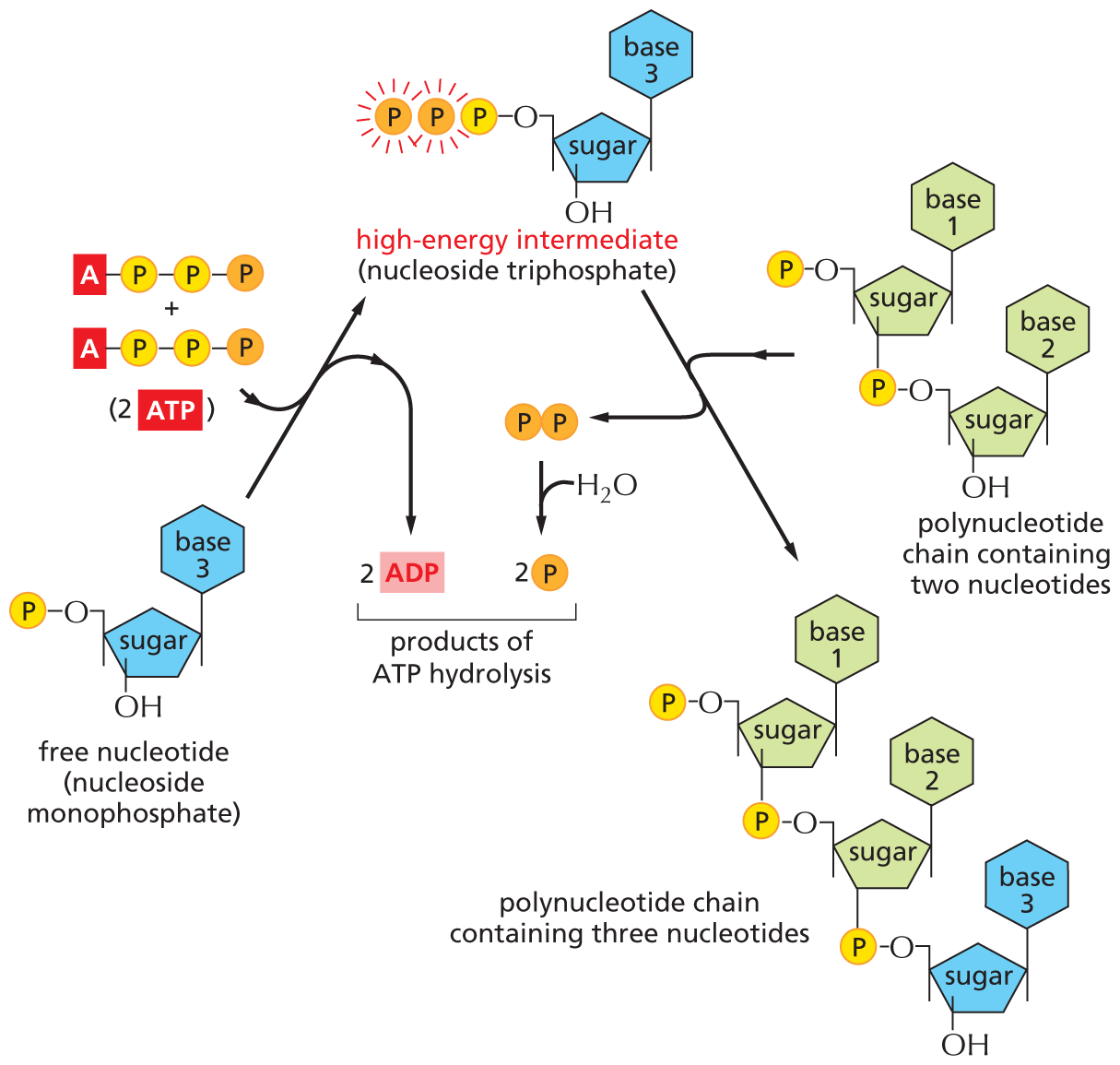
There are limits to what each activated carrier can do in driving biosynthesis. For example, the ΔG for the hydrolysis of ATP to ADP and phosphate depends on the concentrations of all of the reactants, and under the usual conditions in a cell, it is between –46 and –54 kJ/mole. In principle, this hydrolysis reaction can be used to drive an unfavorable reaction with a ΔG of, perhaps, +40 kJ/mole, provided that a suitable reaction path is available. For some biosynthetic reactions, however, even –54 kJ/mole may be insufficient. In these cases, the path of ATP hydrolysis can be altered so that it initially produces AMP and pyrophosphate, a molecule that is itself hydrolyzed in solution in a subsequent step (Figure 3–41). The whole process makes available a total ΔG of about –109 kJ/mole. The biosynthetic reaction involved in the synthesis of nucleic acids (polynucleotides) is driven in this way (Figure 3–42).
More information
A two-part illustration shows the conversion of A T P to A M P and the hydrolysis of pyrophosphate to two phosphate molecules, and a net view of this process. Section A: An illustration shows the conversion of A T P to A M P and the hydrolysis of pyrophosphate to two phosphate molecules.
Section B: An overall view of a reaction is shown, in which A T P is converted to A M P. A water molecule enters the reaction, and the pyrophosphate formed. Pyrophosphate further undergoes a reaction in the presence of water and forms two phosphate groups.
Figure 3–41In an alternative route for the hydrolysis of ATP, pyrophosphate is first formed and then hydrolyzed in solution. This route releases about twice as much free energy as the reaction shown earlier in Figure 3–30. (A) In each of the two successive hydrolysis reactions, an oxygen atom from the participating water molecule is retained in the products, whereas the hydrogen atoms from water form free hydrogen ions, H+. (B) The overall reaction shown in summary form.
QUESTION 3–9
Which of the following reactions will occur only if coupled to a second, energetically favorable reaction?
glucose + O2⟶ CO2 + H2O
CO2 + H2O ⟶ glucose + O2
nucleoside triphosphates ⟶ DNA
nucleotide bases ⟶ nucleoside triphosphates
ADP + phosphate ⟶ ATP
More information
An illustration shows a two step formation of a polynucleotide chain from nucleoside monophosphate via a high energy intermediate. The nucleoside monophosphate has a central cyclopentane sugar ring, in which C 1 is bonded to a base ring at the top and it is numbered 3, C 3 is bonded to a hydroxyl group at the bottom, and C 5 is bonded to an oxygen atom that is further bonded to a phosphate group, represented by a yellow sphere, labeled P. Nucleoside monophosphate undergoes a reaction in the presence of 2 A T P molecules and forms a high-energy intermediate and 2 A D P molecules are released. The high-energy intermediate has a similar structure to that of nucleoside monophosphate, except that it is bonded to a chain of three-phosphate groups. This intermediate undergoes a reaction with a polynucleotide chain containing two nucleotides and during the reaction pyrophosphate is removed, which is further hydrolyzed to form two phosphate groups. The two molecules of A D P and the two phosphate groups are labeled, products of A T P hydrolysis. The structure of a polynucleotide has a cyclopentane sugar ring, in which C 1 is bonded to a base ring at the top and it is numbered 1, C 3 is bonded to a phosphate group at the bottom that further links to the second ring, and C 4 is bonded to a methylene group, which is bonded to an oxygen atom linking to a phosphate group. In the second ring, C 1 is bonded to a base ring at the top and it is numbered 2, C 3 is bonded to a hydroxyl group at the bottom, and C 4 is bonded to a methylene group, which is bonded to an oxygen atom bonded to the linking phosphate group. The product formed, polynucleotide chain containing three nucleotides, has a structure similar to the polynucleotide chain containing two nucleotides, except that the high-energy intermediate is linked to the second ring.
Figure 3–42Synthesis of a polynucleotide, RNA or DNA, is a multistep process driven by ATP hydrolysis. In the first step, a nucleoside monophosphate is activated by the sequential transfer of the terminal phosphate groups from two ATP molecules. The high-energy intermediate formed—a nucleoside triphosphate—exists free in solution until it reacts with the growing end of an RNA or a DNA chain, with release of pyrophosphate. Hydrolysis of the pyrophosphate to phosphate is highly favorable and helps to drive the overall reaction in the direction of polynucleotide synthesis.
ATP will make many appearances throughout this book as a molecule that powers reactions in the cell. In Chapters 13 and 14, we discuss how the cell uses the energy from food to generate ATP. In the next chapter, we learn more about the proteins that make such reactions possible.
A small molecule that stores energy or chemical groups in a form that can be donated to many different metabolic reactions. Examples include ATP, acetyl CoA, and NADH.
Activated carrier that serves as the principal carrier of energy in cells; a nucleoside triphosphate composed of adenine, ribose, and three phosphate groups. (See Figure 2–26.)
Activated carrier that donates the carbon atoms in its readily transferable acetyl group to many metabolic reactions, including the citric acid cycle and fatty acid biosynthesis; the acetyl group is linked to coenzyme A (CoA) by a thioester bond that releases a large amount of energy when hydrolyzed.
A molecule that accepts a hydride ion (H–) from a donor molecule, thereby producing the activated carrier NADH. Widely used in the energy-producing breakdown of sugar molecules. (See Figure 3–34.)
Molecule that accepts a hydride ion (H–) from a donor molecule, thereby producing the activated carrier NADPH; widely used as an electron donor in biosynthetic pathways.