Large amounts of DNA are required to encode all the information needed to make a single-celled bacterium, and far more DNA is needed to encode the information to make a multicellular organism like you. Each human cell contains about 2 meters (m) of DNA; yet the cell nucleus is only 5–8 μm in diameter. Tucking all this material into such a small space is the equivalent of trying to fold 40 km (24 miles) of extremely fine thread into a tennis ball.
In eukaryotic cells, very long, double-stranded DNA molecules are packaged into chromosomes. These chromosomes not only fit handily inside the nucleus, but, after they are duplicated, they can be accurately apportioned between the two daughter cells at each cell division. The complex task of packaging DNA is accomplished by specialized proteins that bind to and fold the DNA, generating a series of loops and coils that provide increasingly higher levels of organization and prevent the DNA from becoming a tangled, unmanageable mess. Amazingly, this DNA is packaged in a way that allows it to remain accessible to all of the enzymes and other proteins that replicate and repair it, and that cause the expression of its genes.
Our discussion of chromosome structure in this chapter will focus entirely on eukaryotic chromosomes. Bacteria typically carry their genes on a single, circular DNA molecule. This molecule is associated with proteins that condense the DNA, but these bacterial proteins differ from the ones that package eukaryotic DNA. Although this bacterial DNA is also called a “chromosome,” its structure differs from that described below.
Eukaryotic DNA Is Packaged into Multiple Chromosomes
In eukaryotes, such as ourselves, nuclear DNA is distributed among a set of different chromosomes. The DNA in a human nucleus, for example, is parceled out into 23 or 24 different types of chromosome, depending on an individual’s sex (males, with their Y chromosome, have an extra type of chromosome that females do not have). Each of these chromosomes consists of a single, enormously long, linear DNA molecule associated with proteins that fold and pack the fine thread of DNA into a more compact structure. This complex of DNA and protein is called chromatin. In addition to the proteins involved in packaging the DNA, chromosomes also associate with many other proteins involved in DNA replication, DNA repair, and gene expression.
With the exception of the gametes (sperm and eggs) and the highly specialized cells that lack DNA entirely (such as mature red blood cells), human cells each contain two copies of every chromosome, one inherited from the mother and one from the father. The maternal and paternal versions of each chromosome are called homologous chromosomes (homologs). The only nonhomologous chromosome pairs in humans are the sex chromosomes in males, where a Y chromosome is inherited from the father and an X chromosome from the mother. (Females inherit one X chromosome from each parent and have no Y chromosome.) Each full set of human chromosomes contains a total of 3.1 × 109 nucleotide pairs of DNA—which together comprise the human genome.
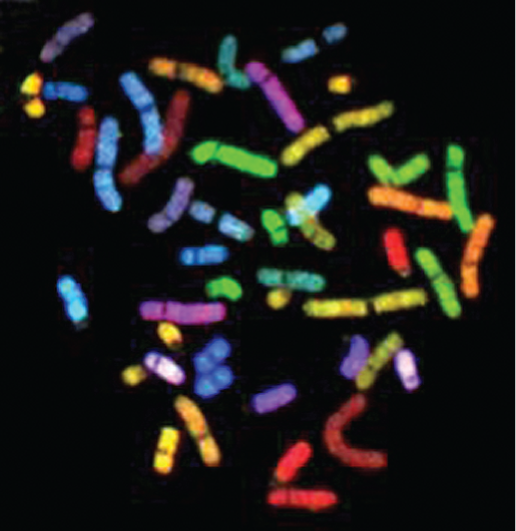
Human chromosomes come in different sizes and can be distinguished from one another using a variety of molecular and microscopic techniques. For example, each chromosome can be “painted” a different color using sets of chromosome-specific, single-stranded DNA molecules coupled to different fluorescent dyes (Figure 5–8A). An earlier and more traditional way of distinguishing one chromosome from another involves staining the chromosomes with dyes that bind to certain types of DNA sequences. These dyes mainly distinguish between DNA that is rich in A-T nucleotide pairs and DNA that is G-C rich, and they produce a predictable pattern of bands along each type of chromosome. The resulting patterns allow each chromosome to be identified and numbered.
More information
A fluorescent micrograph of chromosomes is shown. The Chromosomes are depicted scattered around and out of order, in different colors and ranging in size from 1 to 10 nanometers.
(A)More information
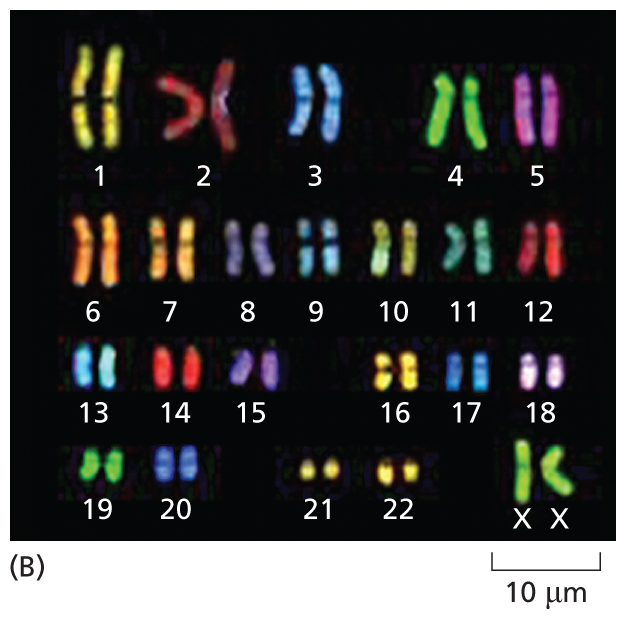
A fluorescent micrograph of chromosomes is shown. In this instance, the chromosomes are shown in pairs and arranged from 1 to number 22 with with another pair labeled X X. The chromosomes range in size from 1 to 10 nanometers.
Figure 5–8Each human chromosome can be “painted” a different color to allow its unambiguous identification. The chromosomes shown here were isolated from a cell undergoing nuclear division (mitosis) and are therefore in a highly compact (condensed) state. Chromosome painting is carried out by exposing the chromosomes to a collection of single-stranded DNA molecules that have been coupled to a combination of fluorescent dyes. For example, single-stranded DNA molecules that match sequences in chromosome 1 are labeled with one specific dye combination, those that match sequences in chromosome 2 with another, and so on. Because the labeled DNA can form base pairs (hybridize) only with its target chromosome (discussed in Chapter 10), each chromosome is differently colored. For such experiments, the chromosomes are treated so that the individual strands of its double-helical DNA partly separate to enable base-pairing with the labeled, single-stranded DNA molecule.
(A) Micrograph showing the array of chromosomes as they originally spilled from the lysed cell. (B) The same chromosomes artificially lined up in their numerical order. This arrangement of the full chromosome set is called a karyotype. (Adapted from N. McNeil and T. Ried, Expert Rev. Mol. Med. 2000:1–14, 2000. With permission from Cambridge University Press.)
An ordered display of the full set of an organism’s chromosomes—such as the 46 chromosomes present in humans—is called a karyotype (Figure 5–8B). If parts of a chromosome are lost, or moved from one chromosome to another, the changes are easy to see in a karyotype. Cytogeneticists analyze karyotypes to detect chromosomal abnormalities that are associated with some inherited disorders (Figure 5–9) and with certain types of cancer (as we see in Chapter 20).
More information
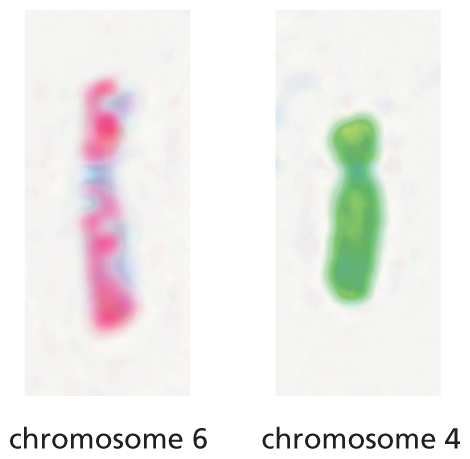
An illustration shows two normal chromosomes: chromosomes 6 and 4. Both the chromosomes are represented as normal by their uniform color.
(A)More information
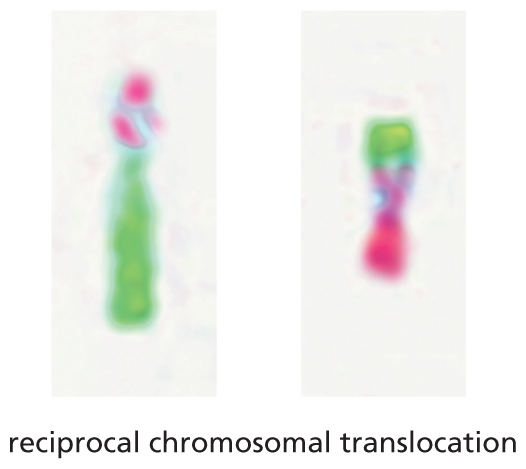
An illustration shows two abnormal chromosomes formed through reciprocal chromosomal translocation. A segment of one chromosome is swapped with a segment of the other chromosome to form two abnormal chromosomes.
(B)Figure 5–9Abnormal chromosomes are associated with some inherited genetic disorders. (A) Two normal human chromosomes, chromosome 6 and chromosome 4, have been subjected to chromosome painting as described in Figure 5–8. (B) In an individual with a reciprocal chromosomal translocation, a segment of one chromosome has been swapped with a segment from the other. Such chromosomal translocations are a frequent event in cancer cells. (Courtesy of Zhenya Tang and the NIGMS Human Genetic Cell Repository at the Coriell Institute for Medical Research.)
Chromosomes Organize and Carry Genetic Information
The most important function of a chromosome is to carry genes—the functional units of heredity. A gene can be defined as a segment of DNA that contains the instructions for making a particular protein or RNA molecule. Most of the RNA molecules encoded by genes are subsequently used to produce a protein. In some cases, however, the RNA molecule is the final product (see Figure 5–7). Like proteins, these RNA molecules have diverse functions in the cell: some serve roles that are structural or catalytic—such as the RNAs that make up the ribosome (discussed in Chapter 7); others play a part in regulating gene expression (discussed in Chapter 8).
More information
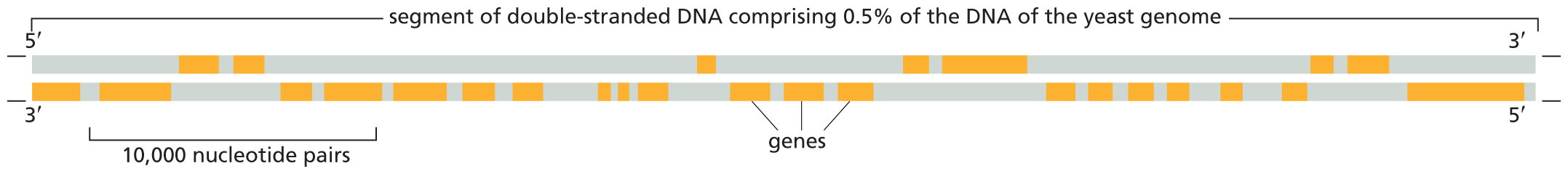
An illustration shows dense gene packing in a yeast chromosome. The D N A comprises a segment of double-stranded D N A comprising 0.5 percent of the D N A of the yeast genome with a 5 to 3 prime upper strand and 3 to 5 prime lower strand. There are numerous genes shown on the D N A that make up more of the D N A than the non-coding regions. A section of 10,000 nucleotide pairs is highlighted and contains five genes.
Figure 5–10In yeast, genes are closely packed along chromosomes. This figure shows a small region of the DNA double helix in one chromosome from the budding yeast S. cerevisiae. The S. cerevisiae genome contains about 12.5 million nucleotide pairs and 6600 genes—spread across 16 chromosomes. Note that, for each gene, only one of the two DNA strands actually encodes the information to make an RNA molecule. This coding region can fall on either strand, as indicated by the orange bars. However, each “gene” is considered to include both the “coding strand” and its complement. The high density of genes is characteristic of S. cerevisiae.
More information
A D N A sequence shows three genes interspersed in large noncoding regions.
The D N A code sequence from row 6 to 9 (CCATGGTGCACCTGACTCCTGAGGAGAAGT; CTGCCGTTACTGCCCTGTGGGGCAAGGTGA; ACGTGGATGAAGTTGGTGGTGAGGCCCTGG; GCAG) is highlighted.
The D N A code sequence from row 13 to 29 (GCTGCTGGTGGTCTAC; CCTTGGACCCAGAGGTTCTTTGAGTCCTTT; GGGGATCTGTCCACTCCTGATGCTGTTATG; GGCAACCCTAAGGTGAAGGCTCATGGCAAG; AAAGTGCTCGGTGCCTTTAGTGATGGCCTG; GCTCACCTGGACAACCTCAAGGGCACCTTT; GCCACACTGAGTGAGCTGCACTGTGACAAG; CTGCACGTGGATCCTGAGAACTTCAGG) is highlighted.
The D N A code sequence from row 49 to 53 (CTCCTGGGCAACGTGCTGGTCTG; TGTGCTGGCCCATCACTTTGGCAAAGAATT; CACCCCACCAGTGCAGGCTGCCTATCAGAA; AGTGGTGGCTGGTGTGGCTAATGCCCTGGC; CCACAAGTATCACTAA) is highlighted.
Figure 5–11In many eukaryotes, genes include an excess of interspersed, noncoding DNA. Presented here is the nucleotide sequence of the human β-globin gene. This gene carries the information that specifies the amino acid sequence of one of the two types of subunits found in hemoglobin, a protein that carries oxygen in the blood. Only the sequence of the coding strand is shown here; the noncoding strand of the double helix carries the complementary sequence. Starting from its 5′ end, such a sequence is read from left to right, like any piece of English text. The segments of the DNA sequence that encode the amino acid sequence of β-globin are highlighted in yellow. We will see in Chapter 7 how this information is transcribed and translated to produce a full-length β-globin protein.
Together, the total genetic information carried by a complete set of the chromosomes present in a cell or organism constitutes its genome. Complete genome sequences have been determined for many thousands of organisms, from E. coli to humans. As might be expected, some correlation exists between the complexity of an organism and the number of genes in its genome. For example, the total number of genes is about 500 for the simplest bacterium and about 25,000 for humans. Bacteria and some single-celled eukaryotes, including the budding yeast S. cerevisiae, have especially compact genomes: the DNA molecules that make up their chromosomes are little more than strings of closely packed genes (Figure 5–10). However, chromosomes from many eukaryotes, including humans, contain—in addition to the coding portions of genes and the specific nucleotide sequences required for normal gene expression—a large excess of extra DNA. Some of this DNA is interspersed between genes, but much is present within them (Figure 5–11). This extra DNA is sometimes called “junk DNA,” because its usefulness to the cell has not been demonstrated. Although this spare DNA does not code for protein, much of it may serve some other biological function. Comparisons of the genome sequences from many different species reveal that small portions of this seemingly superfluous DNA are highly conserved among related species, suggesting their importance for these organisms.
In general, the more complex an organism, the larger is its genome. But this relationship does not always hold. The human genome, for example, is 700 times larger than that of E. coli and 200 times larger than that of S. cerevisiae—but it is 30 times smaller than that of some plants and 10 times smaller than some species of amoeba (see Figure 1–43). Furthermore, how the DNA is apportioned over chromosomes also differs from one species to another. Humans have a total of 46 chromosomes (including both maternal and paternal sets), but a species of small deer has only 7, while some carp species have more than 100. Even closely related species with similar genome sizes can have very different chromosome numbers and sizes (Figure 5–12). Thus, although gene number is roughly correlated with species complexity, there is no simple relationship between gene number, chromosome number, and total genome size. For hundreds of millions of years, the genomes and chromosomes of modern species have each been shaped by a unique history of seemingly random genetic events, acted on by specific selection pressures, as we shall discuss in Chapter 9.
More information
An illustration comparing the chromosomal contents of similar species. The chromosomes of the Chinese muntjac are presented in 22 pairs and a last X Y pair. Each pair is different from the other in size and shape but most are very small. A photo of the Chinese muntjac shows it having brown fur and white horns.
The chromosomes of the Indian muntjac are presented in two pairs and a third set of three. The chromosomes in each set are different from the other sets in size and shape but are generally large. The third set of the Indian muntjac chromosome is labeled as Y 1, X, and Y1. A photo of the Indian muntjac shows it having brown fur and black horns.
Figure 5–12Two closely related species can have similar genome sizes but very different chromosome numbers. In the evolution of the Indian muntjac deer (right), chromosomes that were initially separate, and that remain separate in the Chinese species (left), fused without having a major effect on the number of genes—or the animal. (Left image, courtesy of Deborah Carreno, Natural Wonders Photography; right image, courtesy of Beatrice Bougery.)
Specialized DNA Sequences Are Required for DNA Replication and Chromosome Segregation
To form a functional chromosome, a DNA molecule must do more than simply carry genes: it must be able to be replicated, and the replicated copies must be separated and partitioned equally and reliably into the two daughter cells at each cell division. These processes occur through an ordered series of events, known collectively as the cell cycle. This cycle of cell growth and division is summarized—very briefly—in Figure 5–13; it will be discussed in detail in Chapter 18. Only two broad stages of the cell cycle need concern us in this chapter: interphase, when chromosomes are duplicated, and mitosis, the much briefer stage, when the duplicated chromosomes are distributed, or segregated, to the two daughter nuclei.
More information
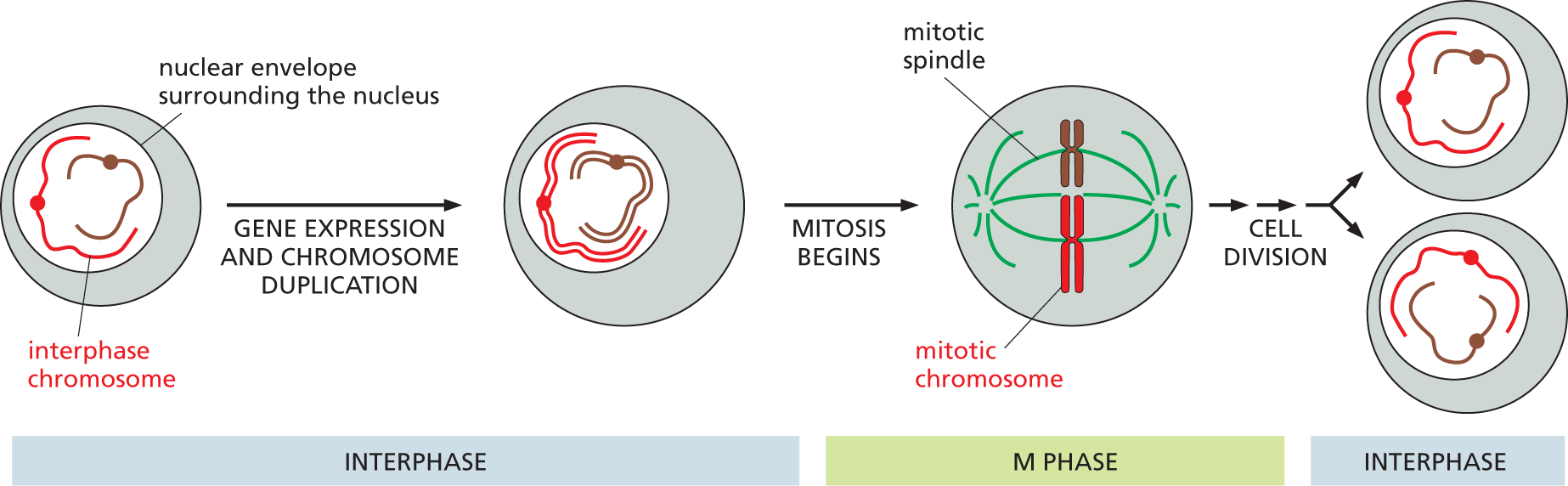
An illustration shows an abbreviated schematic of the cell cycle. The stages are as follows:
Stage 1: interphase: A rounded cell with a nucleus encapsulated within a nuclear envelope contains two interphase chromosomes. This cell undergoes gene expression and chromosome duplication in the first step. The resultant cell has a nucleus containing the duplicated chromosomes.
Stage 2: M phase: The cell containing the duplicated chromosomes undergoes mitosis in the next step to form a mitotic chromosome within the mitotic spindle. The chromosomes are aligned vertically at the center of the mitotic spindle.
Stage 3: Interphase: After mitosis, the cell undergoes cell division to form two daughter cells, each containing a copy of each chromosome within the nucleus.
Figure 5–13The duplication and segregation of chromosomes occur through an ordered cell cycle in proliferating cells. During interphase, the cell expresses many of its genes, and—during part of this phase—it duplicates its chromosomes. Once chromosome duplication is complete, the cell can enter M phase, during which nuclear division, or mitosis, occurs. In mitosis, the duplicated chromosomes condense, gene expression largely ceases, the nuclear envelope breaks down, and the mitotic spindle forms from microtubules and other proteins. The condensed chromosomes are then captured by the mitotic spindle. Next, one complete set of chromosomes is pulled to each end of the cell, and a nuclear envelope forms around each chromosome set. In the final step of M phase, the cell divides to produce two daughter cells. Only two different chromosomes are shown here for simplicity.
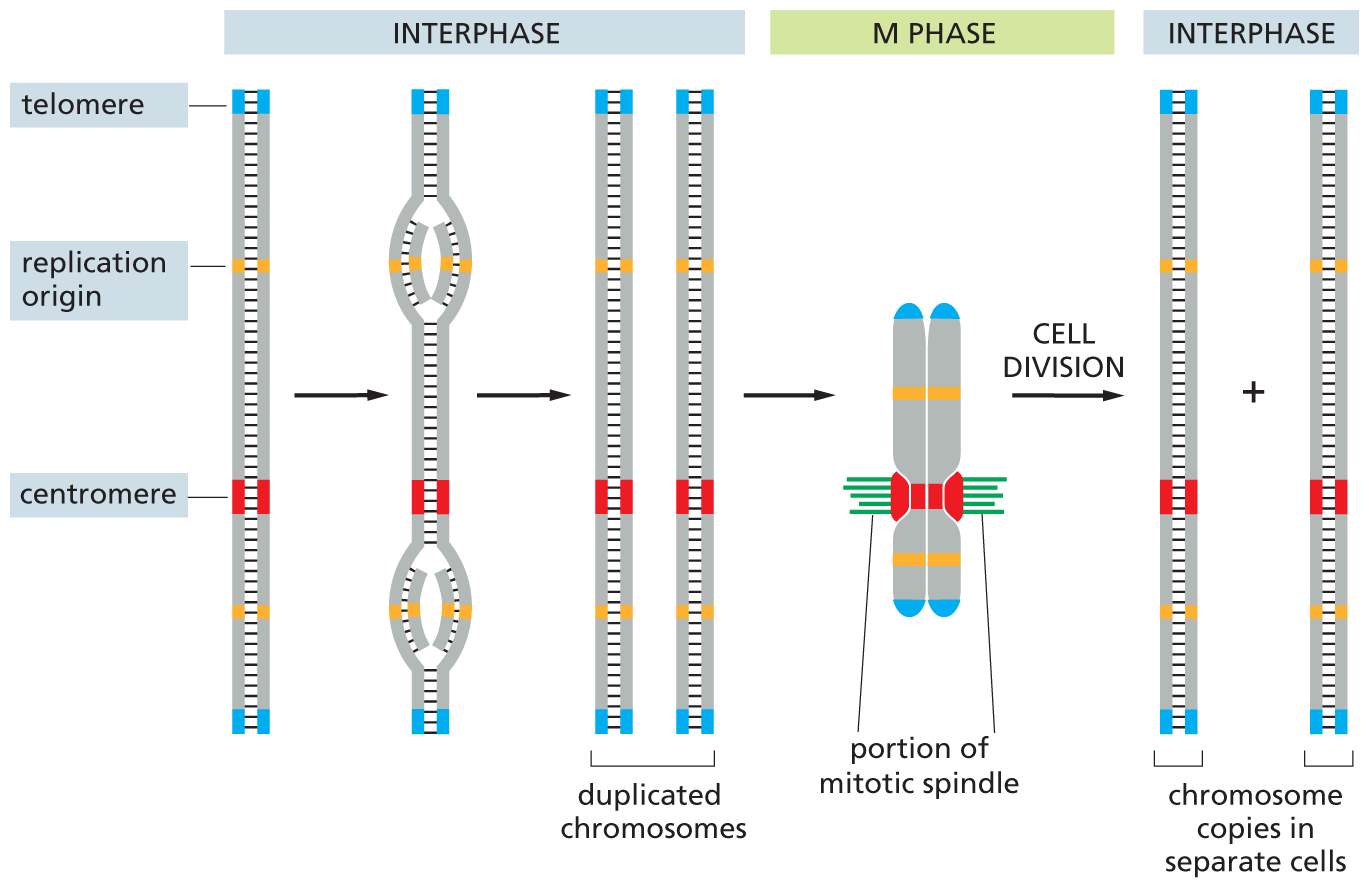
During interphase, chromosomes exist as long, thin threads of DNA in the nucleus and cannot be easily distinguished in the light microscope (see Figure 5–1). We refer to chromosomes in this extended state as interphase chromosomes. It is during interphase that DNA replication takes place. As we discuss in Chapter 6, two specialized DNA sequences, found in all eukaryotes, ensure that this process occurs efficiently. One type of nucleotide sequence, called a replication origin, is the site where DNA replication begins; eukaryotic chromosomes contain many replication origins to allow the long DNA molecules to be replicated rapidly. Another DNA sequence forms the telomeres that mark the ends of each chromosome (Figure 5–14). Telomeres contain repeated nucleotide sequences that are required for the ends of chromosomes to be fully replicated. They also serve as a protective cap that keeps the chromosome tips from being mistaken by the cell as broken DNA in need of repair.
More information
An illustration depicts the D N A sequence elements that are most pertinent to mitosis. The replication is divided into three phases: interphase, M phase, and interphase, respectively. A chromosome is shown as a vertical section of double stranded D N A. There two telomeres, one at the upper end and the other at the lower end. It has a centromere region at three-fourth the height of the strand and two origins of replication, one above and one below the centromere.
Step 1 and step 2 are labeled interphase. In step 1, two bulged curves are formed near the origin of replications, in which two new strands start forming from the inside.
In step 2, a complete duplicated chromosome is formed and that is identical to the first.
In M phase, the two chromosomes are connected at the centromere, and mitotic spindles are shown also attached to the centromere.
In interphase, the two duplicated chromosomes undergo cell division and two chromosome copies separate into two new cells.
Figure 5–14Three DNA sequence elements are needed to produce a eukaryotic chromosome that can be duplicated and then segregated at mitosis. Each chromosome has multiple origins of replication, one centromere, and two telomeres. The sequence of events that a typical chromosome follows during the cell cycle is shown schematically. The DNA replicates in interphase, beginning at the origins of replication and proceeding bidirectionally from each origin along the chromosome out to the telomeres. Telomeres contain DNA sequences that allow for the complete replication of chromosome ends. In M phase, the centromere attaches the compact, duplicated chromosomes to the mitotic spindle so that one copy will be distributed to each daughter cell when the cell divides. Prior to cell division, the centromere also helps to hold the duplicated chromosomes together until they are ready to be pulled apart.
Eukaryotic chromosomes also contain a third type of specialized DNA sequence, called the centromere, that allows duplicated chromosomes to be separated during M phase (see Figure 5–14). During this stage of the cell cycle, the DNA coils up, adopting a more and more compact structure, ultimately forming highly condensed mitotic chromosomes (Figure 5–15). This compact configuration is the state in which the duplicated chromosomes can be most easily visualized (see Figure 5–1). Once the chromosomes have condensed, the centromere allows each duplicated chromosome to attach to the mitotic spindle in a way that directs one copy of each chromosome to be segregated to each of the two daughter cells (see Figure 5–13). We describe the central role that centromeres play in cell division in Chapter 18.
More information
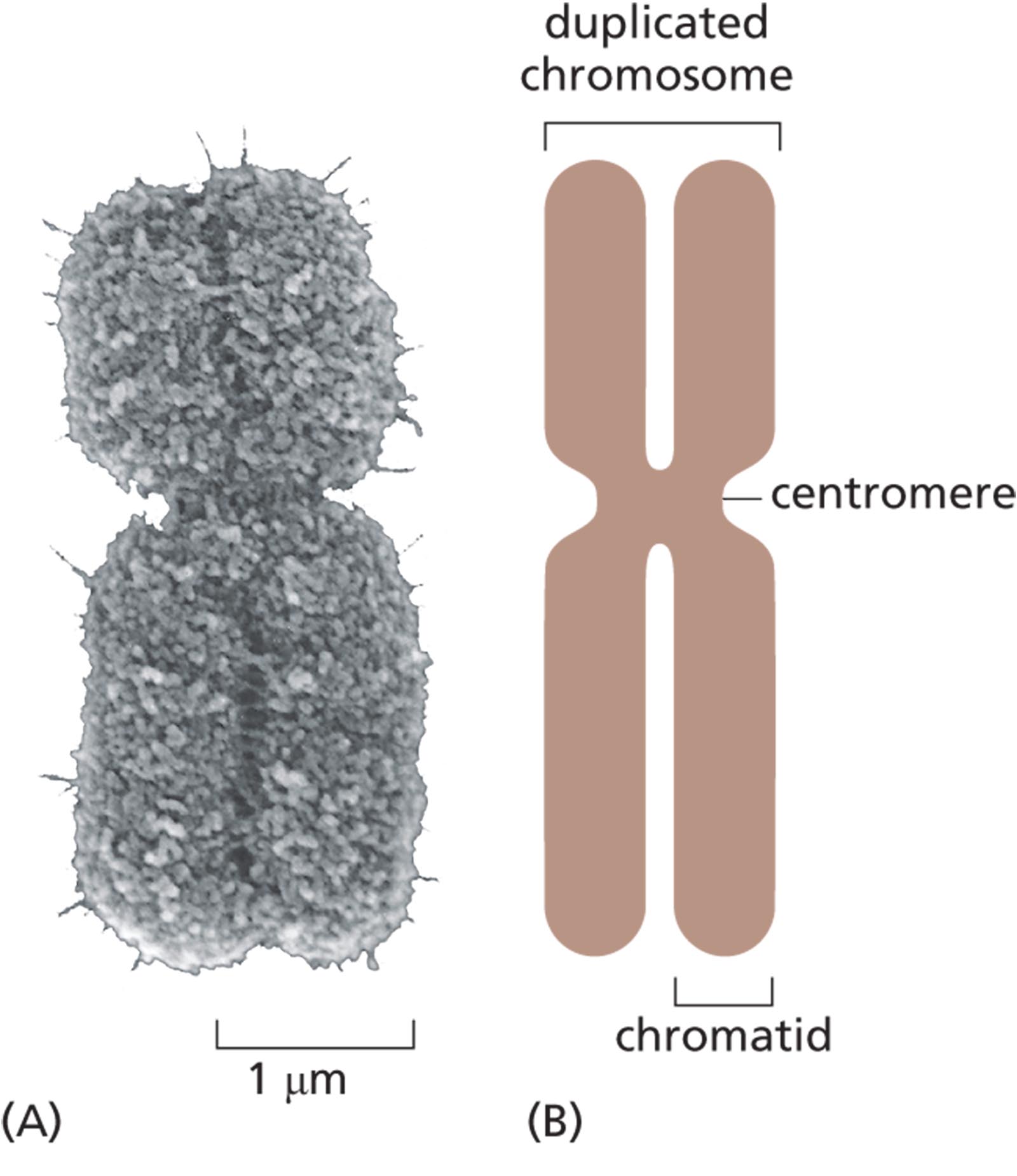
A micrograph shows a duplicated mitotic chromosome that measures about 5 micrometers tall and two micrometers wide. The duplicated chromosome has two identical units, which are closely packed and form an X shape.
An illustration shows the anatomy of an X shaped duplicated chromosome. The constricted mid-section of the chromosome is labeled as centromere, and the one half of the chromosome is labeled as a chromatid. Two combined chromatids form a duplicate chromosome.
Figure 5–15A typical duplicated mitotic chromosome is highly compact. Because DNA is replicated during interphase, each mitotic chromosome contains two identical duplicated DNA molecules (see Figure 5–14). Each of these very long DNA molecules, with its associated proteins, is called a chromatid; as soon as the two sister chromatids separate, they are considered individual chromosomes. (A) A scanning electron micrograph of a mitotic chromosome. The two chromatids are tightly joined together. The constricted region reveals the position of the centromere. (B) A schematic representation of a mitotic chromosome. (A, courtesy of Terry D. Allen.)
Interphase Chromosomes Are Not Randomly Distributed Within the Nucleus
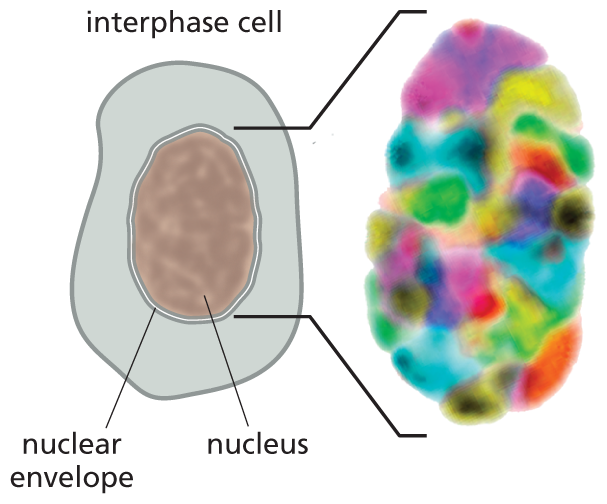
Although interphase chromosomes are much longer and finer than mitotic chromosomes, they are not simply heaped in a jumble inside the nucleus. Instead, they are spatially organized in several ways. First, although interphase chromosomes are constantly undergoing structural and spatial changes in their organization, each tends to occupy a particular region, or territory, of the interphase nucleus (Figure 5–16). This loose organization prevents interphase chromosomes from becoming extensively entangled, like spaghetti in a bowl. In addition, some chromosomal regions are physically attached to particular sites on the nuclear envelope—the pair of concentric membranes that surround the nucleus—or to the underlying nuclear lamina, the protein meshwork that supports the envelope (discussed in Chapter 17). These attachments also help interphase chromosomes remain within their distinct territories.
More information
Section A shows an oval shaped nucleus with a nuclear envelope at the center of an interphase cell. The nucleus occupies a major portion of the cell. An extended view shows the nucleus, made of tightly packed colorful units.
(A)More information
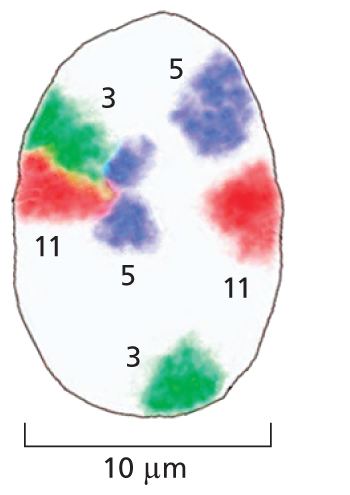
Section B shows a magnified view of the nucleus measuring approximately 10 micrometers in diameter. Three distinct pairs of chromosomes are highlighted in different colors to make the distinction between chromosome 3, 5 and 11. The colorful marking shows that two copies of the same chromosome do not have to be located in the same region of the nucleus.
(B)Figure 5–16Interphase chromosomes occupy their own distinct territories within the nucleus. DNA probes coupled with different fluorescent markers are used to paint individual interphase chromosomes in a human cell. (A) Viewed in a fluorescence microscope, the nucleus is seen to be filled with a patchwork of discrete colors. (B) To highlight their distinct locations, three sets of chromosomes (3, 5, and 11) are singled out. Note that pairs of homologous chromosomes, such as the two copies of chromosome 3 (green), are not generally located in the same position. (Adapted from M.R. Hübner and D.L. Spector, Annu. Rev. Biophys. 39:471–489, 2010.)
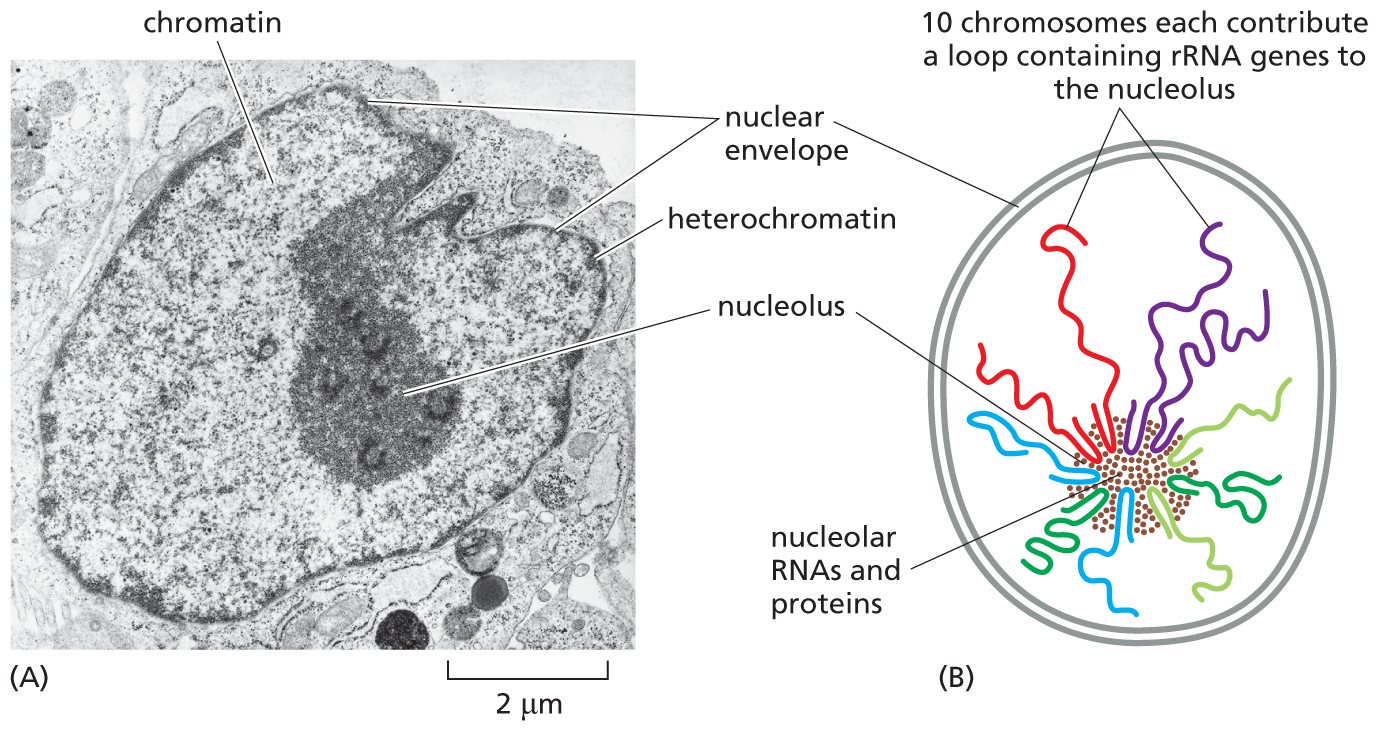
The most easily observable example of chromosomal organization in the interphase nucleus is the nucleolus—a structure large enough to be seen in the light microscope (Figure 5–17A). During interphase, the parts of different chromosomes that carry genes encoding ribosomal RNAs come together to form the nucleolus. In human cells, several hundred copies of these genes are distributed in 10 clusters, located near the tips of five different chromosome pairs (Figure 5–17B). In the nucleolus, ribosomal RNAs are synthesized and combine with proteins to form ribosomes, the cell’s protein-synthesizing machines. As we discuss in Chapter 7, ribosomal RNAs play both structural and catalytic roles in the ribosome.
More information
A micrograph and an illustration show the nucleolus during interphase. A micrograph shows the nucleolus during interphase. The micrograph shows an interphase nucleus, about 6 micrometers in diameter. The interphase nucleus is surrounded by a distinct nuclear envelope and contains a nucleolus about 2 micrometers in diameter. Chromatin and heterochromatin are labeled inside the nucleus and resemble dark, C shapes. A schematic shows the nucleolus during interphase. The schematic shows a rounded nucleus enclosed within a double walled nuclear envelope. The nucleus has the nucleolar R N As and proteins clustered together in the nucleolus. Thin looped strands emanate from the nucleolus. Text corresponding to these thin strands reads, 10 chromosomes each contribute a loop containing r R N A genes to the nucleolus.
Figure 5–17The nucleolus is the most prominent structure in the interphase nucleus. (A) Electron micrograph of a thin section through the nucleus of a human fibroblast. The nucleus is surrounded by the nuclear envelope. Inside the nucleus, the chromatin appears as a diffuse speckled mass; regions that are especially dense are called heterochromatin (dark staining). Heterochromatin contains few genes and is located mainly around the periphery of the nucleus, immediately under the nuclear envelope. The large, dark region within the nucleus is the nucleolus, which contains the genes for ribosomal RNAs. (B) Schematic illustration showing how ribosomal RNA genes, which are clustered near the tips of five different human chromosomes (13, 14, 15, 21, and 22), come together to form the nucleolus, which is a biochemical subcompartment that consists of a dynamic assembly of many macromolecules (see Figure 4–52). These components include ribosomal RNAs (rRNAs) and special proteins, in addition to the indicated DNAs. (A, courtesy of E.G. Jordan and J. McGovern.)
The DNA in Chromosomes Is Highly Condensed
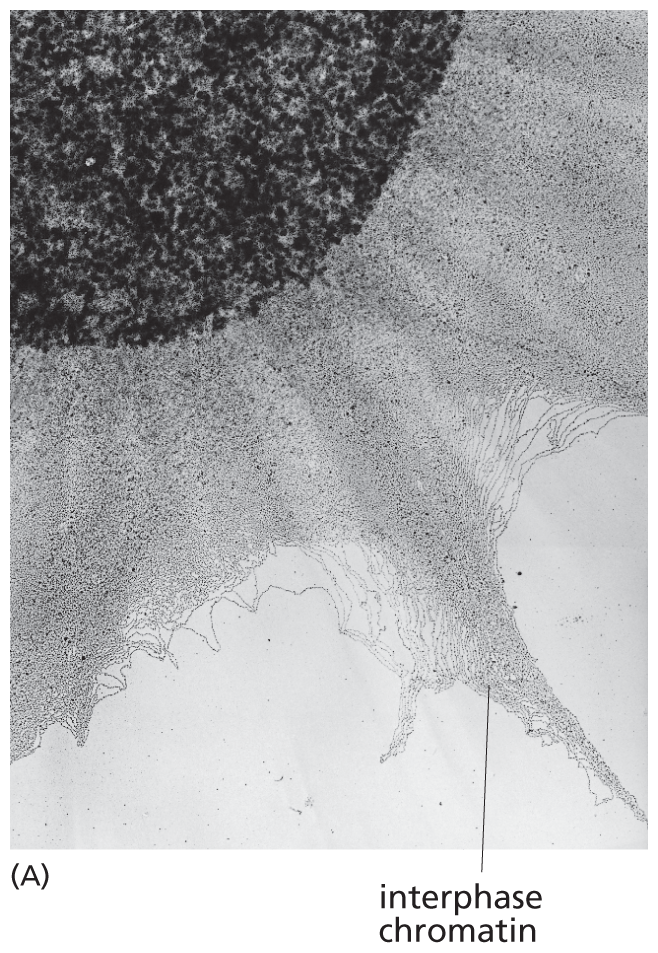
As we have seen, all eukaryotic cells, whether in interphase or mitosis, package their DNA into chromosomes. This packaging involves a good deal of compression. Human chromosome 22, for example, contains about 48 million nucleotide pairs; stretched out end to end, this DNA would extend about 1.5 cm. Yet, during mitosis, chromosome 22 measures only about 2 μm in length—that is, nearly 10,000 times more compact than the DNA would be if it were extended to its full length. Although the DNA of interphase chromosomes is about 10 times less condensed than that of mitotic chromosomes, it is also tightly packed (Figure 5–18). This remarkable feat of condensation is performed by proteins that fold the DNA into higher and higher levels of organization.
More information
Section A shows the nucleus that measures about 10 micrometers in diameter. Extending outward from the nucleus are grey, thin strands of interphase chromatin. They extend about 10 micrometers out from the nucleus.
More information
Section B shows a densely packed mitotic chromosome that resembles the letter X. It measures about 5 micrometers tall and 2 micrometers wide.
Figure 5–18DNA in interphase chromosomes is less compact than in mitotic chromosomes. (A) An electron micrograph showing an enormous tangle of chromatin (DNA with its associated proteins) spilling out of a lysed interphase nucleus. (B) For comparison, a compact, human mitotic chromosome is shown at the same scale. (A, courtesy of Victoria Foe; B, courtesy of Terry D. Allen.)
In the next sections, we introduce the specialized proteins that organize the structure of both interphase and mitotic chromosomes. Bear in mind, though, that this structure is dynamic. Not only do chromosomes condense and decondense through the different phases of the cell cycle, but chromosome packaging must be flexible enough to allow rapid, on-demand access to different regions of the chromosome during interphase, unpacking enough to allow protein complexes to gain access to specific, localized nucleotide sequences for DNA replication, DNA repair, and gene expression.
Nucleosomes Are the Basic Units of Eukaryotic Chromosome Structure
The proteins that bind to DNA to form eukaryotic chromosomes are traditionally divided into two general classes: histones and non-histone chromosomal proteins. Histones are present in enormous quantities (more than 60 million molecules of several different types in each human cell), and their total mass in chromosomes is about equal to that of the DNA itself. Non-histone chromosomal proteins are also present in large numbers that include hundreds of different types. In contrast, there are only a handful of different histone proteins. The complex formed by histone and non-histone chromosomal proteins and nuclear DNA is called chromatin.
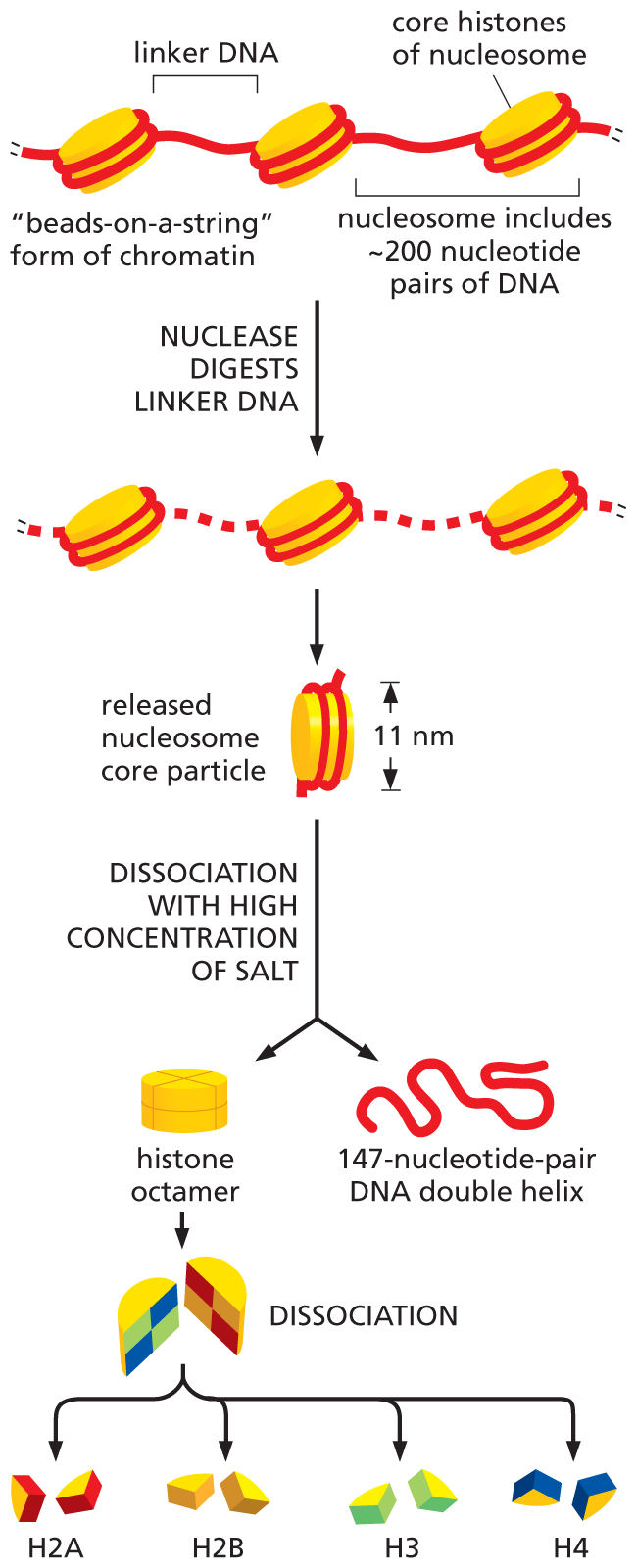
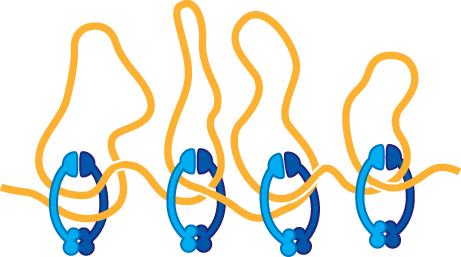
Histones are responsible for the first and most fundamental level of chromatin packing: the formation of the nucleosome. Nucleosomes convert the DNA molecules in an interphase nucleus into a chromatin fiber that is approximately one-third the length of the initial DNA. These chromatin fibers, when examined with an electron microscope, contain clusters of closely packed nucleosomes (Figure 5–19A). If this chromatin is subjected to treatments that cause it to unfold partially, it can then be seen in the electron microscope as a series of “beads on a string” (Figure 5–19B). The string is DNA, and each bead is a nucleosome core particle, which consists of DNA wound around a core of histone proteins.
More information
A micrograph shows packed nucleosomes. They look like a string of densely packed small beads that measure about 5 micrometers in diameter. This string is dark and about 50 micrometers thick.
(A)More information
A micrograph shows decondensed nucleosomes. It resembles a thin string with small 10 micrometer beads on it that are evenly spaced out about 50 micrometers away from each other.
(B)Figure 5–19Nucleosomes can be seen in the electron microscope. (A) Chromatin isolated directly from an interphase nucleus can appear in the electron microscope as a chromatin fiber, composed of packed nucleosomes. (B) Another electron micrograph shows a length of a chromatin fiber that has been experimentally unpacked, or decondensed, after isolation to show the “beads-on-a-string” appearance of the nucleosomes. (A, courtesy of Barbara Hamkalo; B, courtesy of Victoria Foe.)
To determine the structure of the nucleosome core particle, investigators treated chromatin in its unfolded, “beads-on-a-string” form with enzymes called nucleases, which cut the DNA by breaking the phosphodiester bonds between nucleotides. When this nuclease digestion is carried out for a short time, only the exposed DNA between the core particles—the linker DNA—will be cleaved, allowing the core particles to be isolated. An individual nucleosome core particle consists of a complex of eight histone proteins—two molecules each of histones H2A, H2B, H3, and H4—along with a segment of double-stranded DNA, 147 nucleotide pairs long, that winds around this histone octamer (Figure 5–20). The high-resolution structure of the nucleosome core particle revealed, in atomic detail, that the DNA is tightly wrapped around the disc-shaped histone octamer, making 1.7 turns in a left-handed coil (Figure 5–21). The linker DNA between each nucleosome core particle can vary in length from a few nucleotide pairs up to about 80. Technically speaking, a “nucleosome” consists of a nucleosome core particle plus one of its adjacent DNA linkers, as shown in Figure 5–20; however, the term is often used to refer to the nucleosome core particle itself.
More information
An illustration depicts the release of different types of histone octamers due to the dissociation of core histones from linker D N A.
Step 1: The core histones of nucleosomes are arranged like beads-on-a-string, held together by two wrap arounds of linker D N A. The nucleosome includes about 200 nucleotide pairs of D N A. The nuclease digests linker D N A and segments of the linker D N A are missing in the resultant chain such that the cores of histones are no longer connected.
Step 2: A nucleosome core particle of width 11 nanometers is released from the chain and undergoes dissociation with a high concentration of salt to form a histone octamer and a 147-nucleotide-pair D N A double helix.
Step 3: The histone octamer dissociates into H 2 A, H 2 B, H 3, and H 4 which are pairs of proteins that form the histone octamer.
Figure 5–20Nucleosomes contain DNA wrapped around a protein core of eight histone molecules. In a test tube, the nucleosome core particle can be released from chromatin by digestion of the linker DNA with a nuclease, which cleaves the exposed linker DNA but not the DNA wound tightly around the nucleosome core. When the DNA around each isolated nucleosome core particle is released, its length is found to be 147 nucleotide pairs; this DNA wraps around the histone octamer that forms the nucleosome core nearly twice.
More information
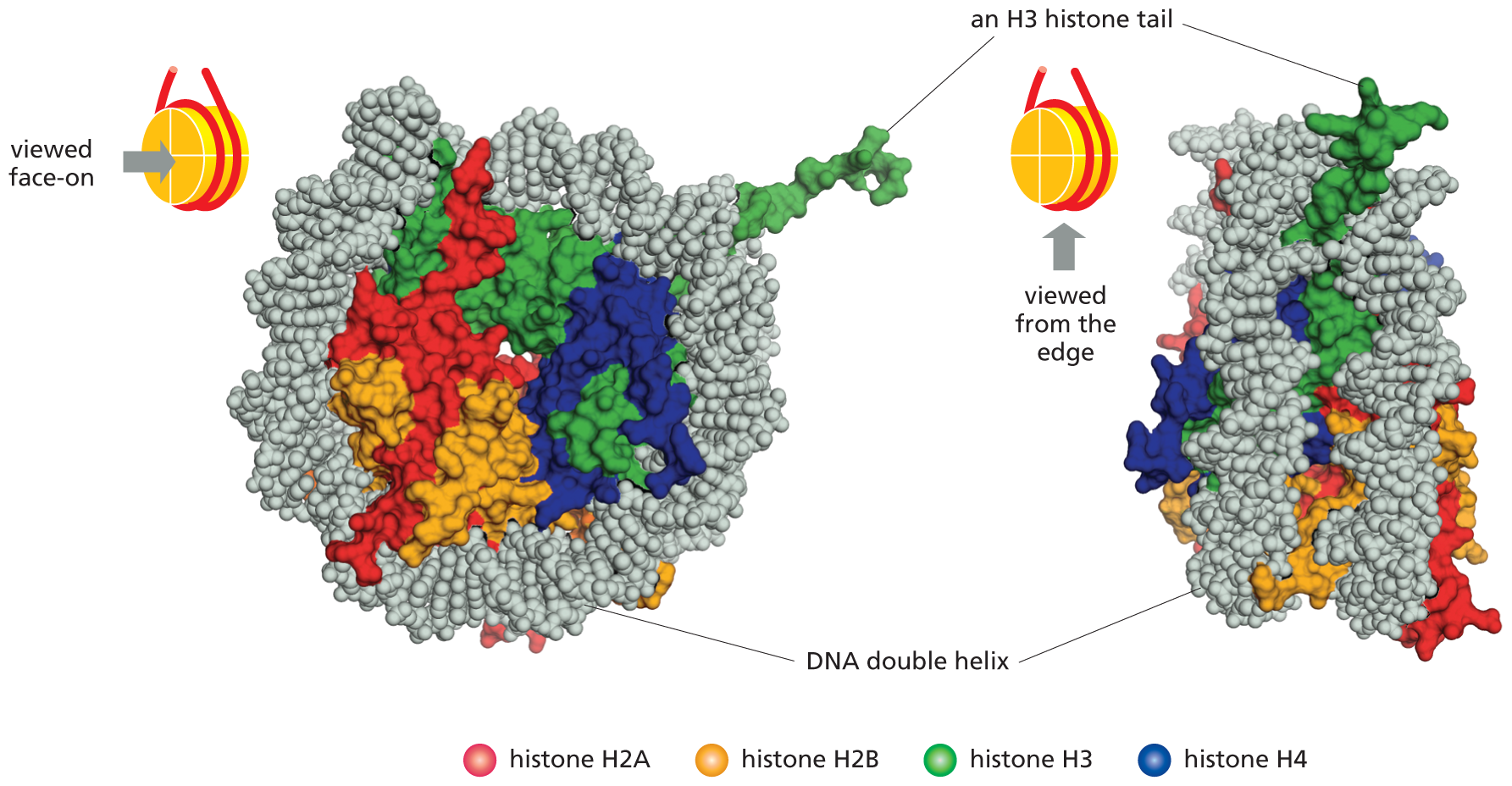
An illustration shows the top view and side view of the structure of the nucleosome determined core particle by using. This model of the histones shows that in the both views, the D N A double helix is wrapped around the 4 histone pairs of proteins that are labeled H 2 A, H 2 B, H 3 and H 4, and are depicted in different colors. In the side view of the histone, the D N A is shown to wrap around the proteins twice and the H 3 histone tail sticks out in between the two wraps.
Figure 5–21The structure of the nucleosome core particle, as determined by x-ray diffraction analysis, reveals how DNA is tightly wrapped around a disc-shaped histone octamer. Two views of a nucleosome core particle are shown here. The two strands of the DNA double helix are shown in gray. A portion of an H3 histone tail (green) can be seen extending from the nucleosome core particle, but the tails of the other histones have been truncated. (From K. Luger et al., Nature 389:251–260, 1997.) (Dynamic Figure) Dynamic molecular simulation shows the spontaneous assembly of a nucleosome from a histone core and a segment of double-stranded DNA. (Courtesy of Stephen E. Farr.)
All four of the histones that make up the octamer are relatively small proteins with a high proportion of positively charged amino acids (lysine and arginine). The positive charges help the histones bind tightly to the negatively charged sugar–phosphate backbone of DNA (see Figure 5–4B). These numerous electrostatic interactions explain in part why DNA of virtually any sequence can bind to a histone octamer. Each of the histones in the octamer also has a long, unstructured N-terminal amino acid “tail” that extends out from the nucleosome core particle (see the H3 tail in Figure 5–21). These histone tails are subject to several types of reversible, covalent chemical modifications that control many aspects of chromatin structure.
The histones that form the nucleosome core are among the most highly conserved of all known eukaryotic proteins: only two differences distinguish the amino acid sequences of histone H4 from peas and from people, for example. This extreme evolutionary conservation reflects the vital role of histones in controlling eukaryotic chromosome structure.
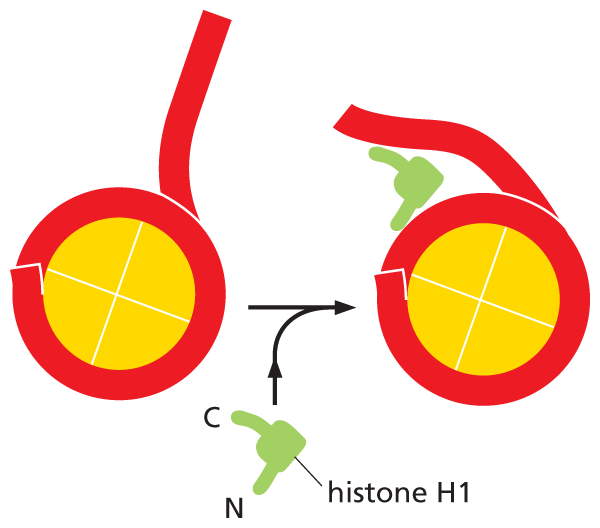
Although long strings of nucleosomes form on most chromosomal DNA, chromatin in the living cell rarely adopts the extended beads-on-a-string form seen in Figure 5–19B. Instead, the nucleosomes are further packed on top of one another to generate a more compact structure, such as the chromatin fiber shown in Figure 5–19A and Movie 5.2. This additional packing of nucleosomes into a chromatin fiber can be aided by a fifth histone called histone H1. This “linker” histone changes the path the DNA takes as it exits the nucleosome core (Figure 5–22). The binding of histone H1 helps to pull adjacent nucleosomes together, producing a more condensed chromatin fiber.
More information
An illustration showing histone H 1 packaging of the nucleosome. H 1 is V shaped and has a C terminus and and N terminus. When a D N A histone complex and H 1 are combined, the long tail of the D N A binds to the H 1 near where it separates from the histone complex which alters the path of the D N A.
Figure 5–22Histone H1 provides additional packaging of nucleosomes in the chromatin fiber. This “linker” histone binds to DNA, altering the path the DNA takes as it exits the nucleosome. In this way, histone H1 helps make the regions of chromatin that it associates with more compact.
Interphase Chromosomes Are Further Organized into Loops by Large Protein Rings
The action of histone proteins—including those that form the nucleosome core particle and the linker histone H1—packages the DNA in an interphase chromosome into a more compact chromatin fiber. But this compression is only the first level of chromosome packaging. The next level converts the linear chromatin fiber into a series of chromatin loops that extend from the center of the chromosome—much like the “bunny ears” formed when we tie a shoelace. These loops allow the interphase chromosome to fold into an even more compact form.
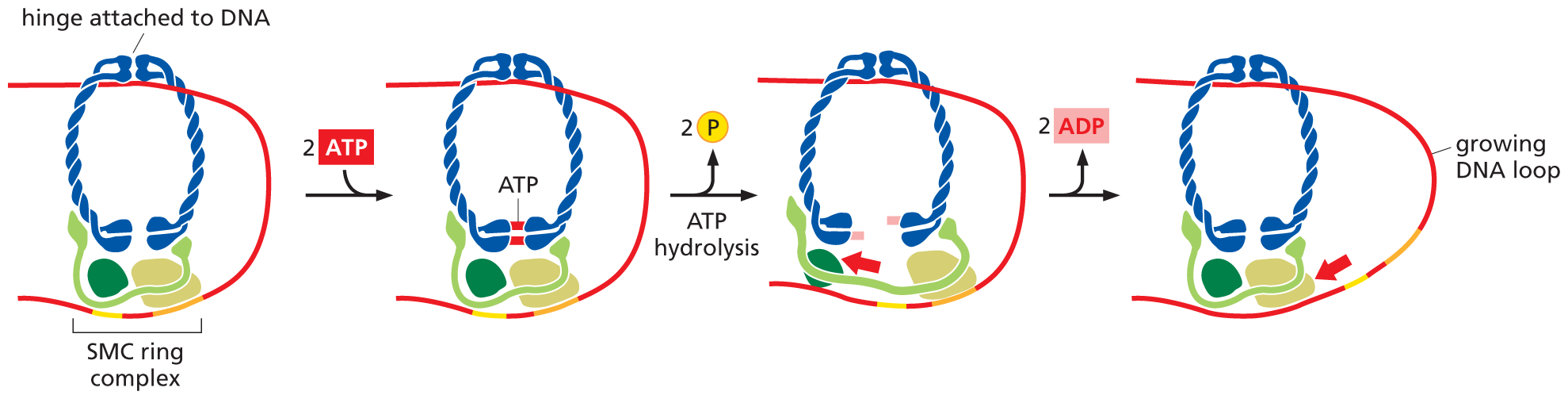
The extrusion of these loops is made possible by a family of proteins that form large rings through which a chromatin fiber can pass. At the heart of this complex is a pair of proteins—the SMC (Structural Maintenance of Chromosomes) proteins—that forms the actual ring. These associate with additional proteins to form an SMC ring complex that uses the energy of ATP hydrolysis to motor along the DNA, pushing out a loop of DNA in its wake (Figure 5–23).
More information
An illustration showing how the S M C complex uses A T P to form chromatin loops. The S M C ring complex consists of a clip-like hinge and three proteins at the bottom holding the ring together. The D N A is attached to the hinge on one side of the ring and the proteins on the other side of the ring. When two molecules of A T P are added to the S M C ring, they bind to the bottom of the hinge keeping the opening in the ring closed. Then the A T P hydrolyze and release 2 phosphate molecules. This causes the ring to open up and the three proteins separate slightly from each other latching on to a new section of D N A. When the ring closes again, the D N A loop between the top and the bottom of the ring grows larger because the D N A was pulled in that direction.
Figure 5–23SMC ring complexes use the energy of ATP hydrolysis to form chromatin loops. Although the detailed mechanism is still uncertain, the SMC complex works like a protein machine that uses energy supplied by ATP hydrolysis to perform this task (see Figure 4–48). In one proposed mechanism, called the “inchworm” model, an SMC ring complex encircles and attaches to a DNA double helix. After hydrolyzing two molecules of ATP, the complex swings opens, widening its “grip” on the DNA. The subsequent release of ADP returns the complex to its original configuration, which brings its “feet” back together a little further along the DNA from where it started. Because the hinge at the top of the complex remains attached to one part of the DNA, this inching movement pushes the DNA at the foot of the complex out into a larger loop. Although the DNA shown here is “naked,” in reality this chromatin fiber would be packaged into nucleosomes, which the SMC ring is large enough to accommodate. The same general mechanism applies to both the SMC ring complexes that organize interphase chromosomes (cohesins) and those that condense mitotic chromosomes (the condensins, to be discussed next). (Dynamic Figure) In the “inchworm” model (below), the activity of an SMC complex extrudes a loop of chromosomal DNA (red).
QUESTION 5–2
Assuming that the histone octamer (shown in Figure 5–20) forms a cylinder 9 nm in diameter and 5 nm in height and that the human genome forms 32 million nucleosomes, what volume of a nucleus that is 6 μm in diameter is occupied by histone octamers? [The volume of a cylinder is πr2h; the volume of a sphere is (4/3)πr3.] What fraction of the total volume of the nucleus do the histone octamers occupy? How does this compare with the volume of the nucleus occupied by human DNA?
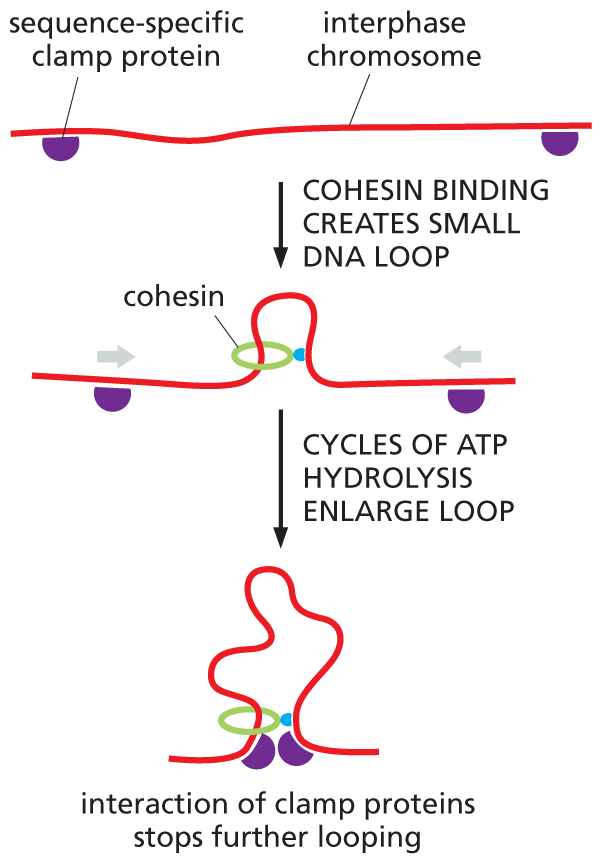
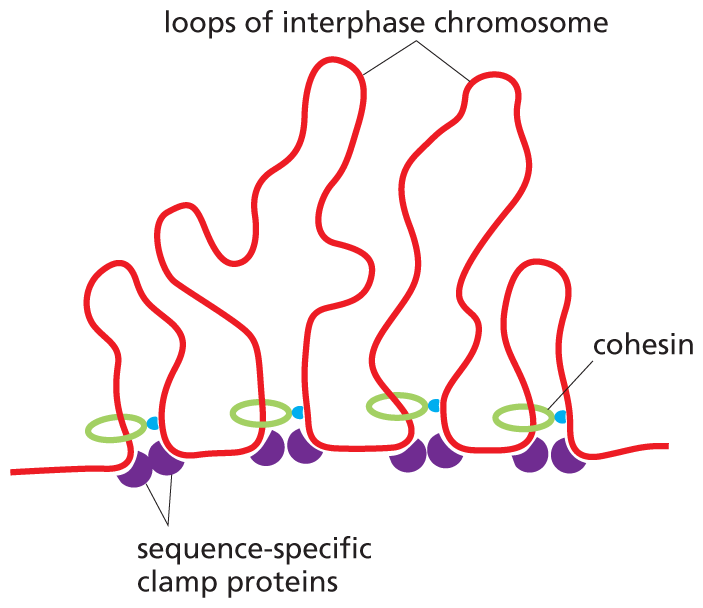
The SMC ring complex that organizes the structure of interphase chromosomes is called cohesin. Multiple cohesins load onto each interphase chromosome, where they produce an extended series of loops. These loops, however, are not formed at random. Cohesin rings will travel along the DNA, extruding loops until they run up against a special sequence-specific clamp protein. These proteins not only stop or stall the moving cohesins, they also bind to one another—an interaction that draws together the DNA at the base of each loop (Figure 5–24). It is the spacing and location of the clamp proteins that dictates the size—and contents—of each chromosomal loop. The resulting structures help to regulate the expression of genes, as we discuss in Chapter 8.
More information
A three step illustration of how chromatin loops are formed. Initially two sequence specific clamp proteins are bound to either end of an interphase chromosome. The clamp proteins are depicted as a small hemisphere on the D N A strand. Then, cohesin binding creates a small D N A loop in the middle of the strand. Cohesin is a circular protein that binds to the D N A and forms a loop that goes through the circular protein. Cycles of A T P hydrolysis enlarge the loop. Once the loop becomes large enough and the clamp proteins are close together and the interaction between the two proteins stops the elongation of the loop.
(A)More information
An illustration showing multiple loops of interphase chromosomes. The D N A sequence is shown with 4 loops in that are irregular shape and of various lengths. The loops have all reached maximum length due to the sequence specific clamp proteins that have reached the cohesin circle and halted elongation.
(B)Figure 5–24Sequence-specific clamp proteins regulate the size of chromatin loops. (A) Cohesins will enlarge chromatin loops until they are stopped by clamp proteins that are bound to specific sequences of DNA. These clamp proteins then interact with one another, drawing together the DNA at the base of each loop. (B) Multiple cohesins operate to divide interphase chromosomes into an extended series of chromatin loops.
In addition to organizing interphase chromosomes, cohesins localized to the centromere play a critical role in holding together the two sister chromatids in a duplicated chromosome (see Figure 5–15). This sister chromatid cohesion, which is maintained through most of mitosis, is required for proper chromosome segregation, as we discuss in Chapter 18.
Chromosomes Undergo an Additional Level of Packing at Mitosis
During mitosis, chromatin becomes so highly condensed that individual chromosomes can be seen in the light microscope (see Figure 5–15A). Although this condensation reduces the length of a typical chromosome only tenfold further, it dramatically alters the chromosome’s appearance and makes the chromosome much easier to segregate into one or other of the daughter cells during cell division. Until recently, the mechanisms that yield this additional level of packing were unknown, but we now understand how a second set of SMC ring proteins, the condensins, plays an important part.
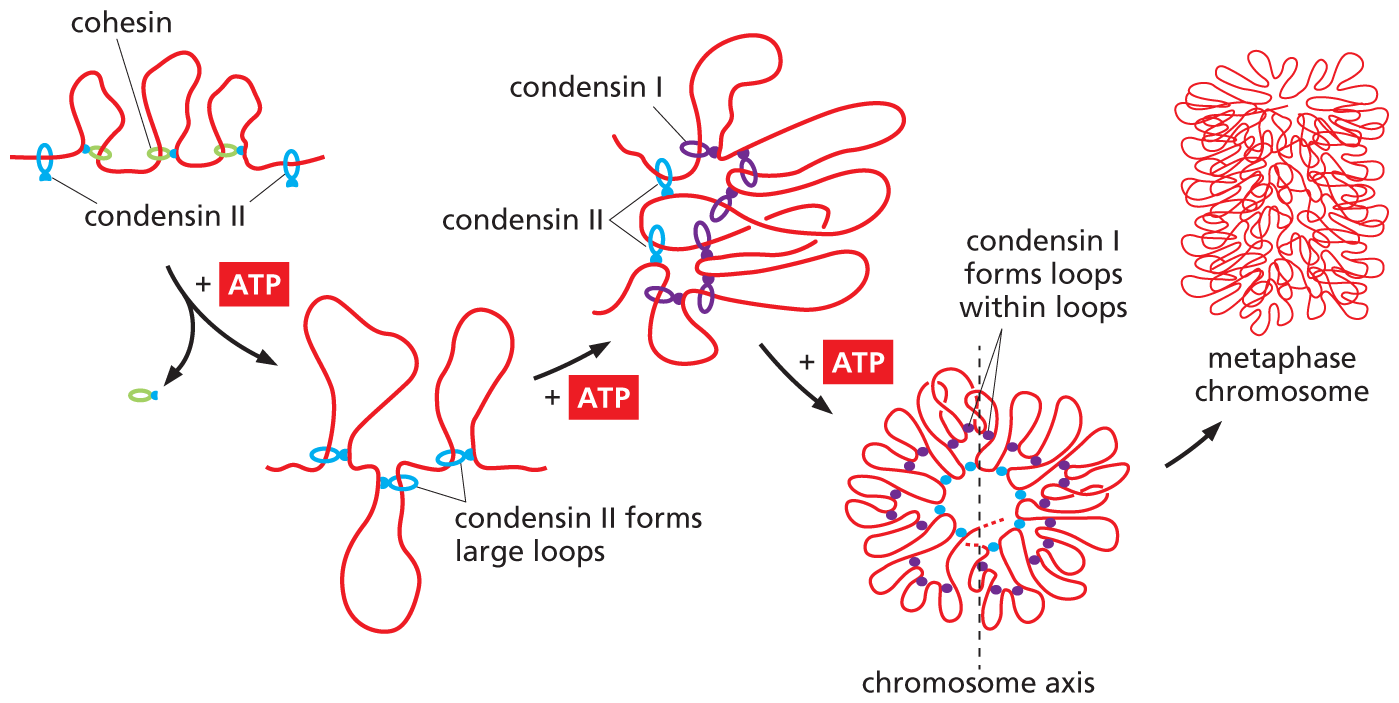
Like their cousin cohesin, condensins are built around ring-shaped SMC proteins. But these complexes contain different SMCs and accessory proteins than do the cohesins. As cells prepare to divide, condensins replace most of the cohesins that formed the loops in the interphase chromosome. These condensins then use the energy of ATP hydrolysis to form loops of their own—including loops within loops—which wind the chromatin into an ever tighter mass of coils (Figure 5–25). It is this final level of condensation that is thought to produce the familiar structure of the mitotic chromosome (Figure 5–26).
More information
An illustration showing how condensin protein forms chromosome loops in metaphase chromosomes. A D N A sequence with three consecutive cohesin loops is shown. The loops have stoped elongating. When condensin II proteins are added to each side of the series of loops, A T P hydrolysis supports formation of larger loops. Text reads: Condensin II forms large loops. With further addition of A T P Condensin II is joined by condensin I protein that further forms loops of groupings of condensin II loops. This starts to orient the D N A in a circular pattern by forming loops within loops. In this circular formation the chromosomal axis runs through the center. This process allows for the condensation of the metaphase chromosome within itself which in this illustration is portrayed as a sequence of looped D N A that is wrapped within itself and stacked on top of itself to form a cylindrical dense chromosome.
Figure 5–25Condensins form loops within loops, folding a mitotic chromosome into a more compact configuration. When cells enter mitosis, most of the cohesins (green) that organized the interphase chromosome are replaced by condensins. Mammalian cells have two condensins: condensin II (blue) forms the initial large chromatin loops and condensin I (purple) then forms a second set of loops inside them. This loops-within-loops organization, combined with the ever-tighter winding of these loops around the chromosome’s central axis, generates the compact structure of the mitotic chromosome. (Adapted from J.H. Gibcus et al., Science 359:eaao6135, 2018.)
More information
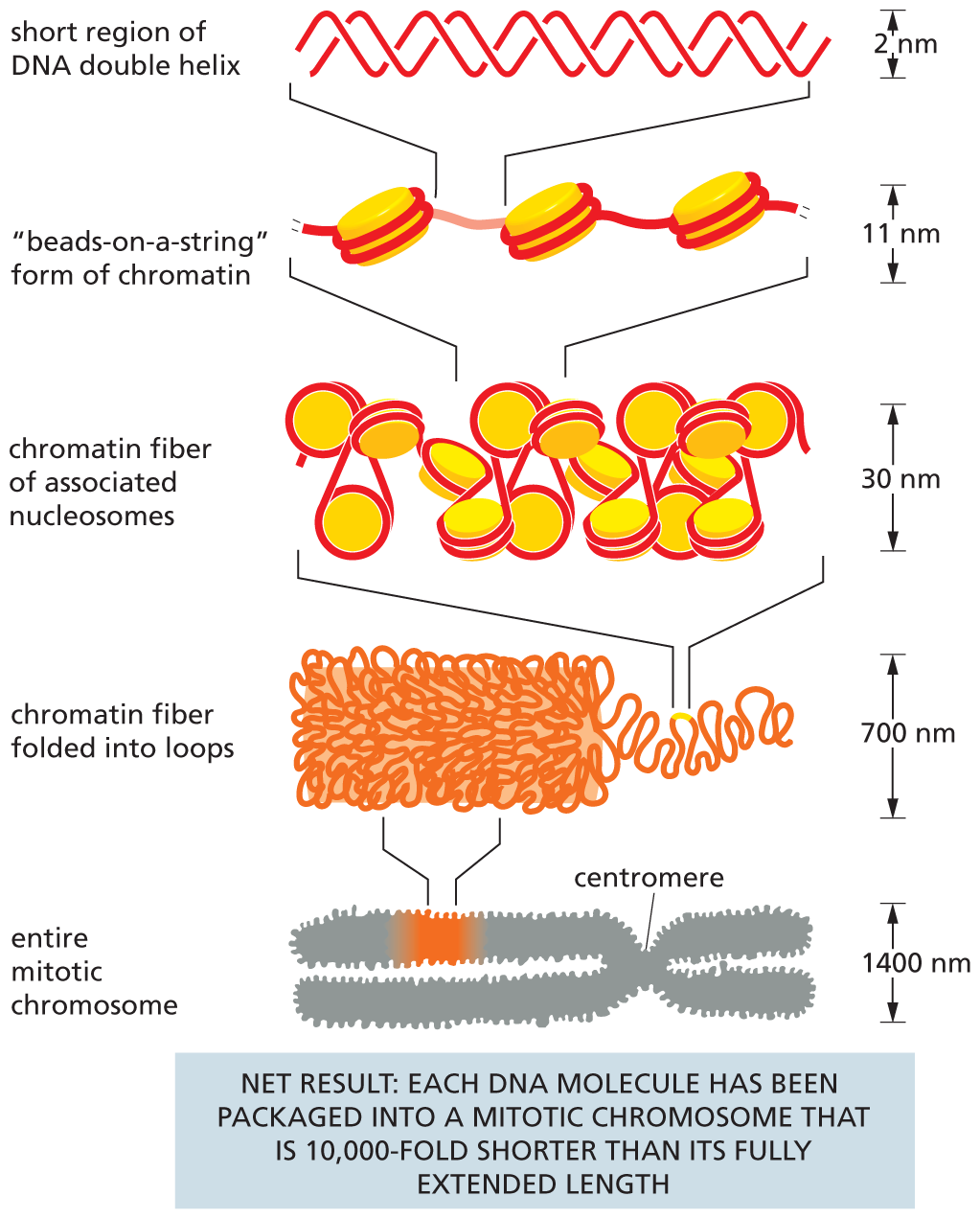
A schematic depicts several levels involved in packing of D N A that results in a D N A molecule packaged into a mitotic chromosome that is 10,000 fold shorter than its fully extended length. Level 1 shows an entire mitotic chromosome. The chromosome has two arms that are attached in the middle. The constricted mid-section of the chromosome is labeled as the centromere. The width of the chromosome is 1400 nanometers. A section of the chromosome is magnified to show level 2.
Level 2 shows a chromatin fiber folded into loops that stack on top of each other. The width of the chain is 700 nanometers. A part of the chromatin fiber is magnified to show level 3.
Level 3 shows chromatin fiber of closely packed nucleosomes. The width of the chain is 30 nanometers. A part of the nucleosome chain is magnified to show level 4.
Level 4 shows core histones of nucleosomes arranged like beads-on-a-string with D N A wrapped around them and linking them. The width of the chain is 11 nanometers. A part of the linker chain connecting the nucleosomes is magnified to show level 5.
Level 5 shows a short region of D N A double helix. The width of the helix is 2 nanometers.
Figure 5–26DNA packing occurs on several levels in chromosomes. This schematic drawing shows the mechanisms thought to give rise to the highly condensed mitotic chromosome. Both histone H1 and a set of specialized non-histone chromosomal proteins are known to help drive these condensations, including chromosome loop-forming clamp proteins and the SMC ring complexes, cohesin and condensin.
Long, threadlike structure composed of DNA and proteins that carries the genetic information of an organism; becomes visible as a distinct entity when a plant or animal cell prepares to divide.
Unit of heredity containing the instructions that dictate the characteristics or phenotype of an organism; in molecular terms, a segment of DNA that directs the production of a particular protein or functional RNA molecule.
The total genetic information carried by all the chromosomes of a cell or organism; in humans, the total number of nucleotide pairs in the 22 autosomes plus the X and Y chromosomes.
Repetitive nucleotide sequence that caps the ends of linear chromosomes. Counteracts the tendency of the chromosome otherwise to shorten with each round of replication.
Specialized DNA sequence that allows duplicated chromosomes to be separated during M phase; can be seen as the constricted region of a mitotic chromosome.
One of a small group of abundant, highly conserved proteins around which DNA wraps to form nucleosomes, structures that represent the most fundamental level of chromatin packing.
Beadlike structural unit of a eukaryotic chromosome composed of a short length of DNA wrapped around an octameric core of histone proteins; includes a nucleosomal core particle (DNA plus histone protein) along with a segment of linker DNA that ties the core particles together.
a large ring-shaped structure that contains two Structural Maintenance of Chromosomes (SMC) protein subunits at its core; the ring these proteins form is large enough for nucleosomes to pass easily through it. Cohesins and condensins are two major types.
a ring-shaped SMC protein complex that organizes interphase chromosomes into a long series of large chromatin loops; in addition, a special subset of cohesins holds together the sister chromatids after DNA has been replicated.