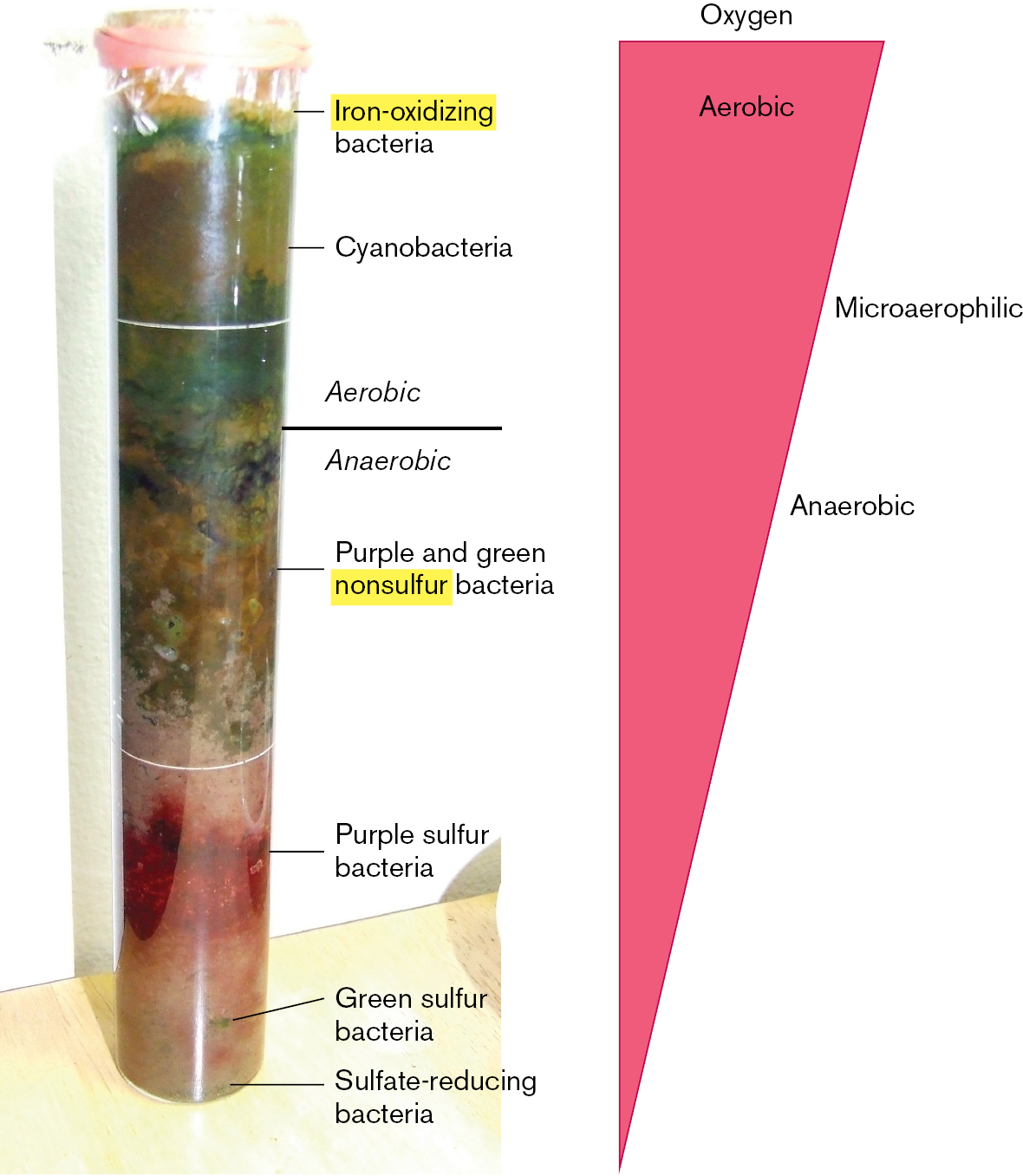
Koch’s growth of microbes in pure culture founded the systematic study of microbial physiology and biochemistry. But how does pure culture relate to “natural” environments, such as a forest or a human intestine, where countless kinds of microbes interact with each other and with multicellular hosts? The opposite of pure culture is the Winogradsky column (Fig. 1.24). The Russian microbiologist Sergei Winogradsky (1856–1953) devised this model wetland ecosystem containing regions of enrichment for microbes of diverse metabolism.
More information
A two-part illustration shows the Winogradsky column, a wetland model ecosystem.
A photo of the Winogradsky column. The model consists of a thin plastic column, in which differently colored layers are visible. The top section of the column is labeled Aerobic. The growth in this section is green, a few different shades are visible. The middle section of the column is labeled Anaerobic. The growth in this section is green and brown. The bottom section of the column is labeled Anaerobic. The growth in this section is red and yellow-green. From the top of the column to the bottom, the bacteria growing are labeled, iron-oxidizing bacteria, cyanobacteria, purple and green nonsulfur bacteria, purple sulfur bacteria, green sulfur bacteria and sulfate reducing bacteria.
An inverted triangle describing oxygen availability within the Winogradsky column. From top to bottom, the triangle is labeled Aerobic, Micoaerophilic, and Anaerobic. The whole triangle is labeled oxygen. The largest section of the triangle is at the top and the smallest section is at the bottom, correlating to the amount of oxygen moving from the top to the bottom of the Winogradsky column.
FIGURE 1.24 ■Winogradsky column. A wetland model ecosystem designed by Sergei Winogradsky.PHOTO BY TIMOTHY A. WILKERSON
A typical Winogradsky column consists of a glass tube containing mud (a source of wetland bacteria) mixed with shredded newsprint (an organic carbon source) and calcium salts of sulfate and carbonate (an inorganic carbon source for autotrophs). After exposure to light for several weeks, several zones of color develop, full of mineral-metabolizing bacteria. At the top, cyanobacteria conduct photosynthesis, using light energy to split water and produce molecular oxygen. Below, purple sulfur bacteria use photosynthesis to split hydrogen sulfide, producing sulfur. At the bottom, with O2 exhausted, bacteria reduce (donate electrons to) alternative electron acceptors such as sulfate. Sulfate-reducing bacteria produce hydrogen sulfide and precipitate iron.
In hindsight, the invention of pure culture eclipsed the equally important study of microbial ecology (discussed in Chapters 21 and 22). Microbes cycle the many minerals essential for all life, including all atmospheric nitrogen gas and much of the oxygen. Yet, less than 0.1% of all microbial species can be cultured in the laboratory. In natural environments, uncultured microbes make up the majority of Earth’s entire biosphere. Only the outer skin of Earth supports complex multicellular life. The depths of Earth’s crust, to at least 3 kilometers (km) down, as well as the atmosphere 15 km out into the stratosphere, remain the domain of microbes. So, to a first approximation, Earth’s ecology is microbial ecology.
Environmental Microbes Support Ecosystems
The first microbiologists to culture microbes in the laboratory selected the kinds of nutrients that feed humans, such as beef broth or potatoes. Winogradsky, however, suspected that other kinds of microbes living in soil or wetlands consume more exotic fare. Soil samples were known to oxidize hydrogen gas, and this activity was eliminated by treatment with heat or acid, suggesting microbial origin. Ammonia in sewage was oxidized by donating electrons to oxygen, forming nitrate. Nitrate formation was eliminated by antibacterial treatment. These findings suggested the existence of microbes that “eat” hydrogen gas or ammonia instead of beef or potatoes, but no one could isolate these microbes in culture.
Winogradsky waded through marshes to discover microbes with metabolisms quite alien from human digestion. For example, he discovered that species of the bacterium Beggiatoa oxidize hydrogen sulfide (H2S) to sulfuric acid (H2SO4). Beggiatoa fixes carbon dioxide into biomass without consuming any organic food. Organisms that feed solely on inorganic minerals are known as chemolithotrophs, or lithotrophs (discussed further in Chapters 4 and 14).
The lithotrophs studied by Winogradsky could not be grown on Koch’s plate media containing agar or gelatin. The bacteria that Winogradsky isolated can grow only on inorganic minerals; in fact, some species are actually poisoned by organic food. For example, nitrifiers convert ammonia to nitrate, forming a crucial part of the nitrogen cycle in natural ecosystems. Winogradsky cultured nitrifiers on a totally inorganic solution containing ammonia and silica gel, which supported no other kind of organism. This experiment was an early example of enrichment culture, the use of selective growth media that support certain classes of microbial metabolism while excluding others.
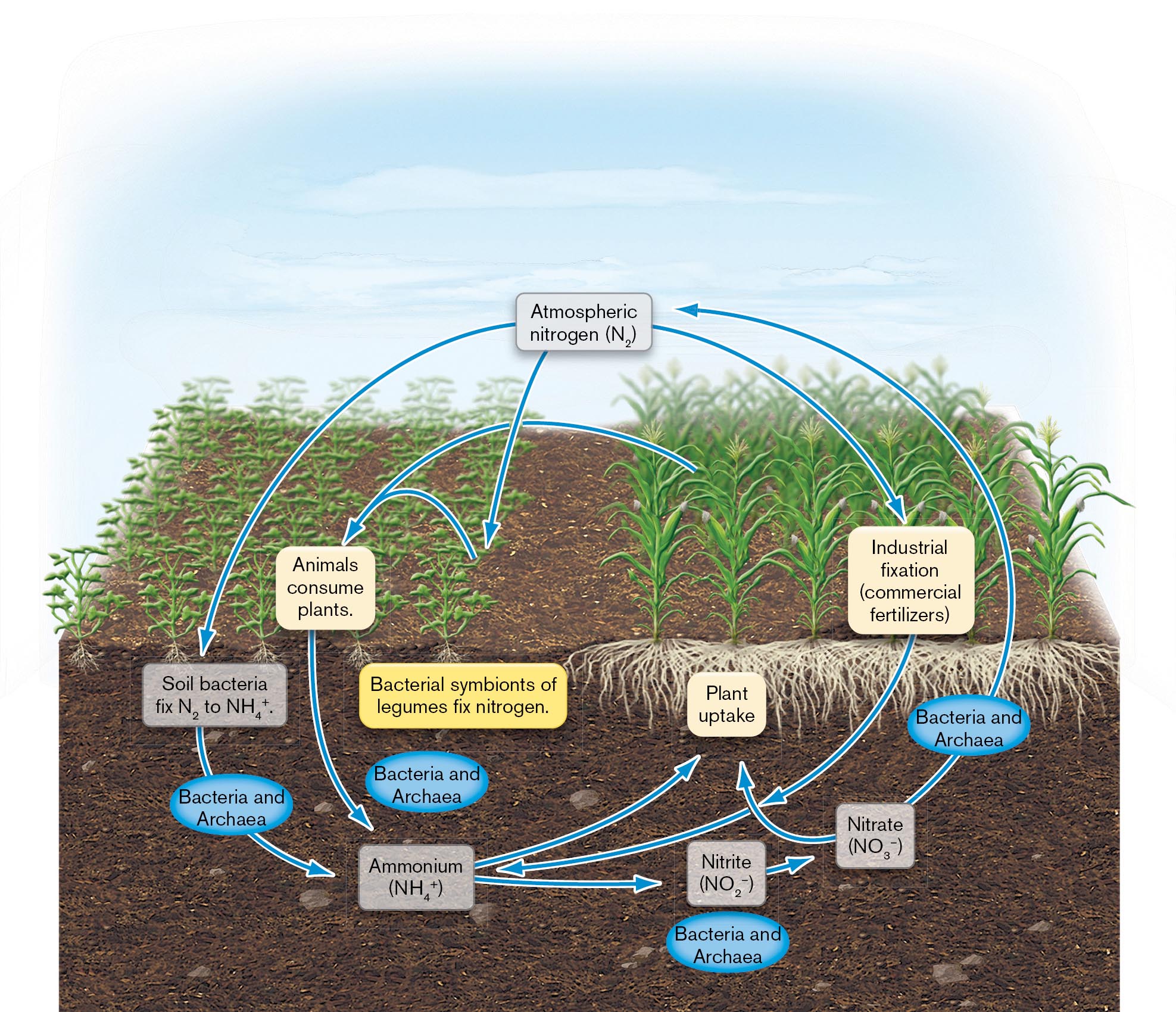
Later microbial ecologists showed that bacteria perform unique roles in geochemical cycling, the global interconversion of inorganic and organic forms of nitrogen, sulfur, phosphorus, and other minerals. Without these essential conversions (nutrient cycles), no plants or animals could live. Bacteria and archaea fix nitrogen (N2) by reducing it to ammonia (NH3), the form of nitrogen assimilated by plants. This process is called nitrogen fixation (Fig. 1.25). Other bacterial species oxidize ammonium ions (NH4+) in several stages back to nitrogen gas. These microbial cycles are heavily affected by human technology leading to climate change (discussed further in Chapters 21 and 22).
More information
An illustration of the process of the global nitrogen cycle. The nitrogen cycle begins from atmospheric nitrogen, N subscript 2, which can be fixed by soil bacteria to ammonium, N H subscript 4 plus, or atmospheric nitrogen can undergo industrial fixation, such as through commercial fertilizers, to ammonium. Ammonium can be taken up by plants or converted by bacteria and archaea into nitrite, N O subscript 2 minus. Nitrite is further converted to nitrate, N O subscript 3 minus, by bacteria and archaea. Nitrate can be taken up by plants or it can be used by bacteria and archaea and released as atmospheric nitrogen. Plants that uptake ammonium, can be eaten by animals, which will eventually return the nitrogen to ammonium in the earth. Atmospheric nitrogen can also be fixed by bacterial symbionts of legumes.
FIGURE 1.25 ■The global nitrogen cycle. All life depends on these oxidative and reductive conversions of nitrogen—most of which are performed only by microbes.
Thought Question
1.8 Why don’t all living organisms fix their own nitrogen? Consider the structure of a dinitrogen molecule, N≡N.
ANSWER ANSWER
Nitrogen fixation requires a tremendous amount of energy, about 30 molecules of ATP per dinitrogen molecule converted to ammonia (discussed in Chapter 15). In a community containing adequate nitrogen sources, organisms that lose the nitrogen fixation pathway make more efficient use of their energy reserves than do those that spend energy to fix nitrogen from the atmosphere. Another consideration is that nitrogenase is an oxygen-sensitive enzyme, whereas plants, animals, and fungi are aerobes. In order to fix nitrogen, aerobic organisms need to develop complex mechanisms to keep oxygen away from nitrogenase.
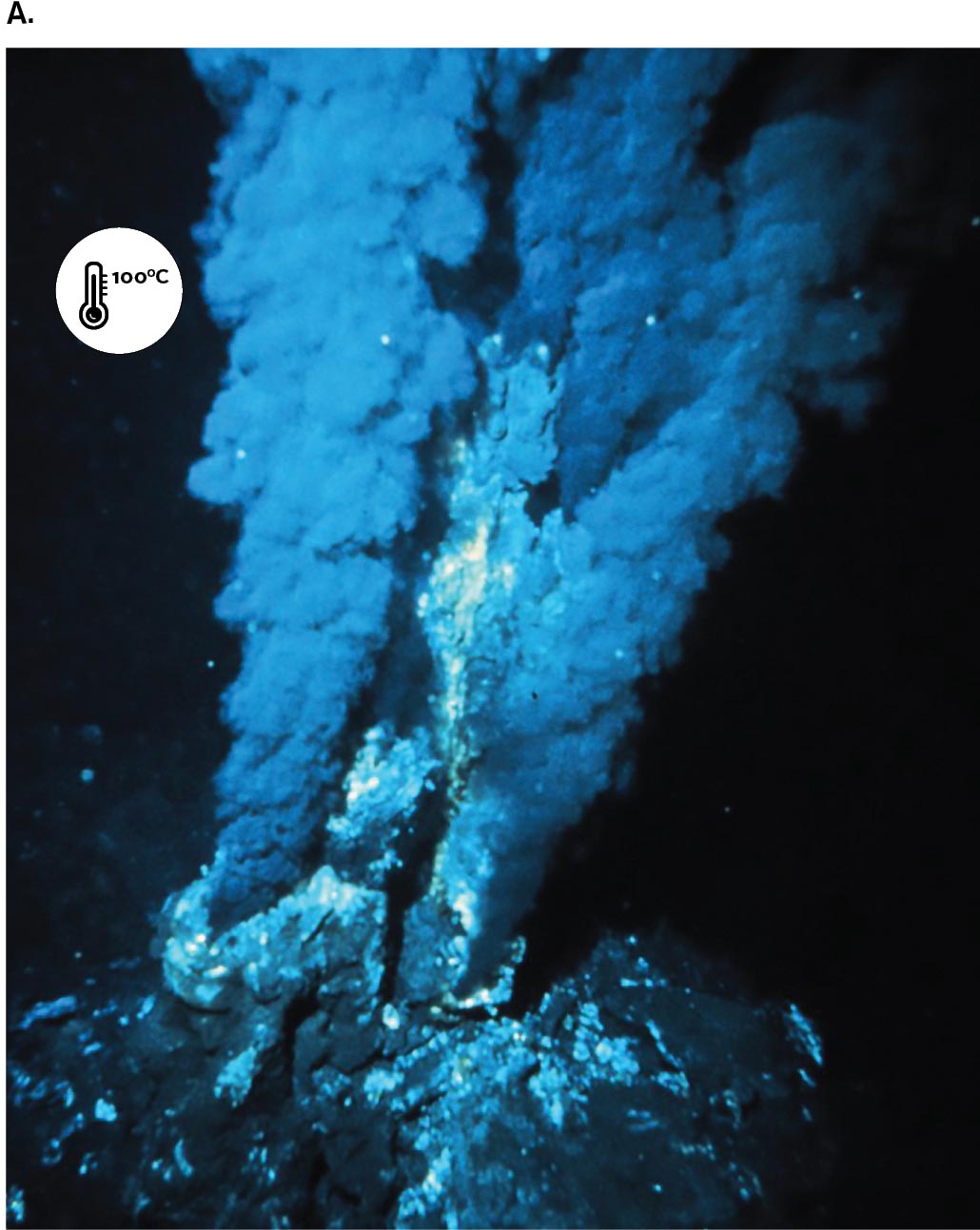
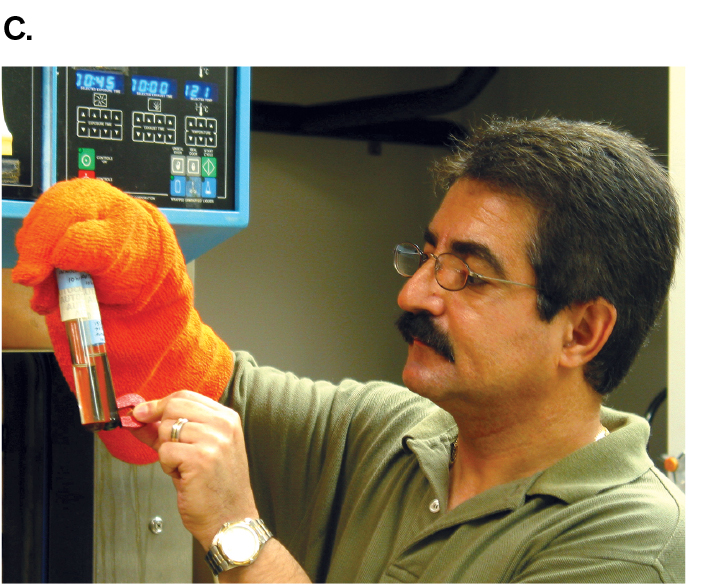
Today, microbes with unusual properties, such as the ability to digest toxic wastes or withstand extreme temperatures, have valuable applications in industry and bioremediation. For this reason, microbial ecology is a priority for funding by the National Science Foundation (NSF). From 1998 to 2004, the NSF was directed by marine microbial ecologist Rita Colwell (Colwell’s research is described in Chapter 21). At NSF, Colwell founded the Biocomplexity Initiative to study complex interactions between microbes and other life in the environment. Such research includes the discovery of extremophiles, microbes from environments with extreme heat, salinity, or acidity. For example, hydrothermal vents in the ocean floor spew superheated water and minerals that feed hyperthermophiles (Fig. 1.26A). A record-setting hyperthermophile is the archaeon Geogemma barossii (Fig. 1.26B), isolated by microbiologist Kazem Kashefi from the Mothra vent field named for the fictional monster. G. barossii reduces rust (iron oxide; Fe2O3) to the magnetic mineral magnetite (Fe3O4) while growing in an autoclave at 121°C (250°F), a temperature high enough to kill all other known organisms. A simple magnet can be used to show that Geogemma converts nonmagnetic Fe2O3 to the magnetic Fe3O4 (Fig. 1.26C).
More information
Three images related to the thermophilic archaeon Geogemma barossii are shown. Image A is a photograph of an ocean hydrothermal vent. Image B is a micrograph of Geogemma and iron oxide. Image C is photograph of a scientist holding a sample tube of Geogemma.
Image A is a photograph of sulfide plumes rising from an ocean hydrothermal vent. It is indicated that the sulfide plumes are a temperature of 100 degrees Celsius.
More information
Image B is an electron micrograph showing Geogemma strain 121, and iron oxide. Geogemma is a large round archaeon with many thin flagella protruding from a single point. Geogemma has a diameter of 1.5 micrometers. Next to Geogemma, there is a buildup of iron oxide.
More information
Image C is a photograph of microbiologist Kazem Kashefi holding a magnet to a sample tube of Geogemma strain 121. Kashefi is wearing a thick, orange, heat resistant glove to hold the sample tube.
FIGURE 1.26 ■An extreme thermophile reduces iron oxide to magnetite.A. Hydrothermal vent at ocean floor. B.Geogemma barossii is a round archaeon with a tuft of flagella (transmission electron micrograph). C. Kazem Kashefi, now at Michigan State University, pulls a live culture of “strain 121” (Geogemma) out of an autoclave generally used to kill all living organisms at 121°C (250°F). OAR/NURP/NOAAK. KASHEFI AND D. R. LOVLEY. 2003. SCIENCE 301:934KAZEM KASHEFI/MICHIGAN STATE UNIVERSITY
Microbial Endosymbiosis with Plants and Animals
The pure-culture model of microbiology, a powerful tool of discovery, nonetheless disregarded the fact that outside the laboratory, all microbes live in the presence of other kinds of life. Many live in endosymbiosis with multicellular organisms. Endosymbiosis is the partnership of a host organism with its associated endosymbionts, microbes that grow within a host body or within a host cell. The first person to describe endosymbiosis was Martinus Beijerinck, who observed nitrogen-fixing bacteria called rhizobia (singular, rhizobium) within the cells of plants. Rhizobia induce the roots of legumes such as soybean plants to form special nodules to fix nitrogen into biomass, which is shared with the plant cells.
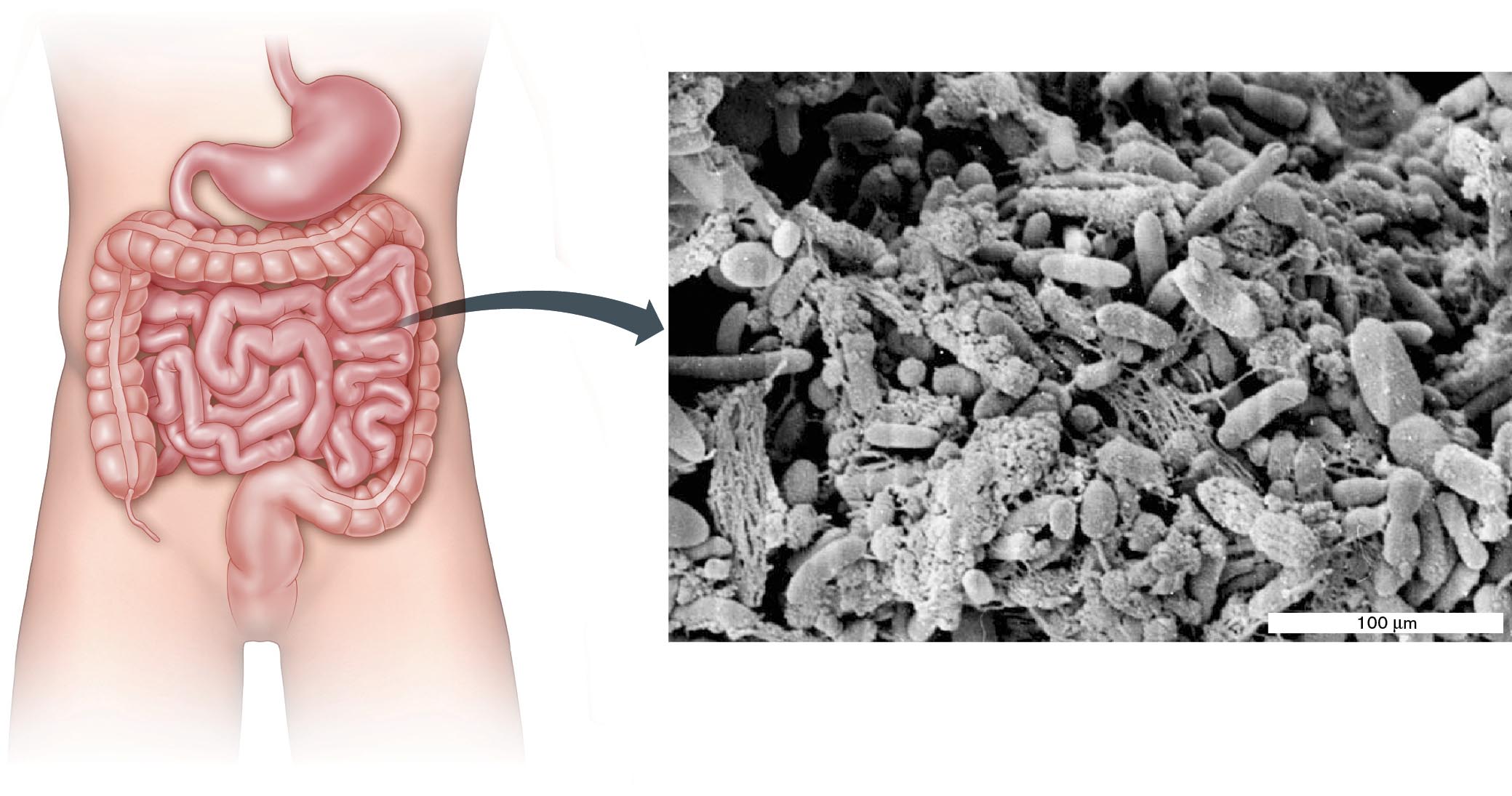
Microbial endosymbiosis occurs everywhere. Endosymbiotic microbes make essential nutritional contributions to host animals. Invertebrates such as hydras and corals harbor endosymbiotic phototrophs that provide products of photosynthesis in return for protection and nutrients. Other endosymbionts produce antibiotics or toxins that thwart predators. Among vertebrates, ruminant animals such as cattle, as well as insects such as termites, require digestive bacteria such as Bacteroides to break down cellulose and other plant polymers. Even humans obtain about 15% of our nutrition from bacteria growing within the colon (Fig. 1.27).
More information
A diagram of a human digestive tract connected to an enlarged electron micrograph of a bacterial biofilm. The micrograph shows stringy pieces of food with a variety of bacterial species surrounding them. Rod-shaped bacteria and cocci are visible. Most species are less than 100 micrometers in length or diameter.
A diagram of a human digestive tract connected to an enlarged electron micrograph of a bacterial biofilm. The micrograph shows stringy pieces of food with a variety of bacterial species surrounding them. Rod-shaped bacteria and cocci are visible. Most species are less than 100 micrometers in length or diameter.
FIGURE 1.27 ■Intestinal microbiota form a biofilm.Blowup: Bacterial biofilm on the surface of a residual food particle.AMERICAN SOCIETY FOR MICROBIOLOGYMACFARLANE, S., ET AL. 2005. J. APPL. ENVIRON. MICROBIOL.71:7483
Some intestinal bacteria, such as Escherichia coli, grow as biofilms, organized multispecies communities adhering to a surface—in this case the surface of colonic epithelial cells. Biofilms play major roles in all ecosystems and within parts of the human body (discussed in Chapters 4, 13, and 21). The biofilm shown magnified in Figure 1.27 is attached to the surface of a digested food particle.
Today we know that all multicellular organisms possess a microbiota, or microbiome, the collection of all microbes associated with an organism or habitat. Remarkably, our concept of “multicellular organism” has changed to include its microbiome as a functional part of the organism. Physicians consider the human microbiome to be a part of the body, as essential as a limb or an organ. Bacteria that normally inhabit the human intestine and skin protect our bodies from infection by pathogens. Gut bacteria regulate the development of our immune system and even send signals to the brain (discussed in Chapter 13). In 2016, the U.S. government announced the National Microbiome Initiative to advance understanding of how microbiomes contribute to our health and the environment. Microbiomes are discussed further in Chapters 21 and 23.
Note: The term “microbiota” refers to the ecological community of microbes living within or upon an organism, such as the human body. “Microbiome” refers to the community of microbes associated with an organism or with a different defined habitat, such as soil or plants; the term emphasizes the microbes’ collective DNA sequences. Despite these subtle distinctions, we use the terms interchangeably in this book. Another term, “metagenome,” refers specifically to the collective DNA sequences found in all the microbes of a microbiome.
Thought Question
1.9 Could endosymbiosis occur today; that is, could a small microbe be engulfed by a larger one and evolve into an endosymbiont, and then into an organelle? Explain.
ANSWER ANSWER
There are many examples today of endosymbiotic associations that look like evolution of an interdependent relationship. For example, a paramecium can acquire internalized chlorella algae that conduct photosynthesis and provide nutrients for the protozoan host. However, in the dark, where light is unavailable, the paramecium may instead digest the chlorella for food. Other bacteria, such as Wolbachia species, have evolved as endosymbionts of insect cells. In some cases, the insect absolutely requires bacterial endosymbionts to provide amino acids but digests the endosymbiont when the nutrients are no longer needed. Still other insects host permanent endosymbiotic bacteria such as Buchnera, which are transmitted vertically (from parent to offspring). These permanent endosymbionts may be on their way to evolving into organelles.
Microbial Life on Other Planets
The abundance of life on Earth—and the evidence of life’s appearance early in Earth’s history—leads us to ask whether microbial life has emerged on other planets. Molecules that spontaneously formed in Stanley Miller’s and Joan Oró’s experiments are also found in meteorites and comets. This observation led Oró to propose that the first chemicals of life could have come from outer space, perhaps carried by comets. Furthermore, at the time that life arose on Earth, Earth’s geochemistry resembled that of other planets, such as Mars. In 2012, to seek evidence for Martian life, NASA landed the Mars Science Laboratory, or Curiosity rover, near the base of a mountain on the planet Mars (Fig. 1.28). The car-sized rover has a laser to drill into rock, X-ray and fluorescence analyzers, and camera microscopes. As of this writing, Curiosity continues its mission, testing the Martian soil for water, organic compounds, and other potential evidence of microbial life. The question became more interesting in 2018 when the orbital radar detector MARSIS found evidence of water buried beneath Mars’s southern ice cap—similar to ice-buried lakes that support microbial life on Earth.
More information
A photo of the Mars Curiosity rover on Mars.
FIGURE 1.28 ■Mars Curiosity rover. The rover explores Mars for evidence of microbial life.NASA/JPL-CALTECH/MSSS
To Summarize
Sergei Winogradsky developed the first system of enrichment culture, called the Winogradsky column, to grow microbes from natural environments.
Chemolithotrophs (or lithotrophs) metabolize inorganic minerals, such as ammonia, instead of the organic nutrients used by the microbes isolated by Koch.
Geochemical cycling depends on bacteria and archaea that cycle nitrogen, phosphorus, and other minerals throughout the biosphere.
Endosymbionts are microbes that live within host organisms and may provide essential functions for their hosts, such as nitrogen fixation, digestion of food molecules, or protection from predation.
Martinus Beijerinck was the first to demonstrate that nitrogen-fixing rhizobia grow as endosymbionts within leguminous plants.
The microbiome, or microbial community, of a multicellular host is now considered a functional part of the host organism.
NASA’s Curiosity rover now explores the planet Mars for signs of microbial life outside Earth.
A glass tube containing a stratified environment that causes specific microbes to grow at particular levels; a type of enrichment culture for the growth of microbes from wetland environments.
The metabolic ability to absorb and convert solar energy into chemical energy for biosynthesis. Autotrophic photosynthesis, or photoautotrophy, includes CO2 fixation.
An organism that grows only in an extreme environment; that is, an environment including one or more conditions that are “extreme” relative to the conditions for human life.
 ANSWER
ANSWER ANSWER
ANSWER