MEMORY AND THE EQUIPOTENTIALITY DEBATE
As Wernicke and others were demonstrating the power of the new localization theories, a partially contrasting line of evidence regarding the brain’s storage of memory was developing, and came to prominence in the United States in the early 1900s. In 1902 a young American psychologist, Shepherd Ivory Franz (1874–1933), published a study of the effects of cortical ablations on cats that had previously been trained to escape from a “puzzle box.”12 This study was very much in the tradition of Flourens—except that instead of looking at the generalized effects of ablation as Flourens had, Franz was interested in the effects on a specific, learned response. His innovation was to combine ablation with animal training.

Figure 3.8 Karl Lashley (1890–1958).
Figure 3.8 Karl Lashley (1890–1958).
If the memories for the learned responses were localized in specific small regions of the cortex, then only those ablations involving those particular regions should have affected the responses. Franz’s study, however, found localization only of a highly general sort, in that lesions of the frontal cortex caused the responses to be lost, while lesions elsewhere did not. Even more significant, it seemed to Franz, was the fact that the frontally ablated animals were sometimes able to relearn the escape response quite easily and quickly. Obviously then, if the removed regions had been somehow responsible for the original learning, altogether different brain parts were able to perform highly similar functions in the relearning. This finding led Franz to distrust the strong localization theory, and to recall and respect Flourens’s old dictum that the brain functions as a relatively undifferentiated whole.
Franz spent much of his subsequent career in hospital settings and remained highly impressed by this plasticity and flexibility of the brain. He observed that human patients suffering with lost functionality from localized brain damage sometimes managed to reacquire those functions, either partially or completely. The recovery was likely to be greatest in young patients. Infants and children with extensive left-side damage, for example, regularly acquired language functions normally even though their Broca’s and Wernicke’s areas had been destroyed. Franz concluded, “Everything tended to show that there are not the definite and exact functions for parts of the cerebrum which were posited [by the most enthusiastic localizationists], but that there is rather a possibility of substitution.”13
In 1915, Franz gained an ally and colleague for further animal research—Karl Spencer Lashley (1890–1958; Figure 3.8). Although his Ph.D. was in genetics, Lashley had been attracted to psychology through his friendship and collaboration at Johns Hopkins University with John B. Watson, the charismatic founder of the behaviorist movement (see Chapter 9). Watson and the behaviorists ruled out any experimental methods involving introspection or reports of conscious states, and concentrated on the strictly observable and “objective” behaviors of their subjects. Animals such as tame white rats were favored subjects because they could be easily observed, their environments could be controlled, and they obviously could not mislead the experimenters with unverifiable subjective reports. Typical experiments studied the ability of animals to learn how to solve such problems as the running of mazes. They demonstrated that hungry white rats could gradually learn to solve complicated mazes if rewarded with food whenever their trial-and-error explorations brought them to the ends of the mazes. The number of false turns or “errors” tended to decrease with each succeeding trial, thereby providing objective measures of the animals’ learning ability.
When Lashley met Franz, the two decided to pool their expertise. Lashley trained white rats on a maze and another simple learning task, and Franz performed selective brain ablations. As they expected, given Franz’s earlier work, they found little experimental evidence for localization, and they said so in a pair of papers in 1917. Subsequently Lashley, on his own, now trained by Franz on ablation techniques, conducted a more extensive series of studies. He trained large numbers of white rats on a wide variety of mazes and other tasks, followed by systematically varying ablations.
His highly detailed findings, recounted in the classic 1929 book Brain Mechanisms and Intelligence and summarized by the graph in Figure 3.9, seemed to firmly refute the localization-of-memory hypothesis. The graph shows the numbers of errors made by rats running mazes of three levels of difficulty, following six degrees of ablation. On the whole, the specific locations of the ablations made little difference at all. Of much greater importance were absolute sizes of the ablations, and the difficulty levels of the mazes involved. In general, large ablations impeded performance on all mazes more than small ablations did, and the effect was much more marked on the difficult mazes than the easy ones. These results indicated that memories seemed to be contained within the entire cortex, and not selectively localized in small parts of it.
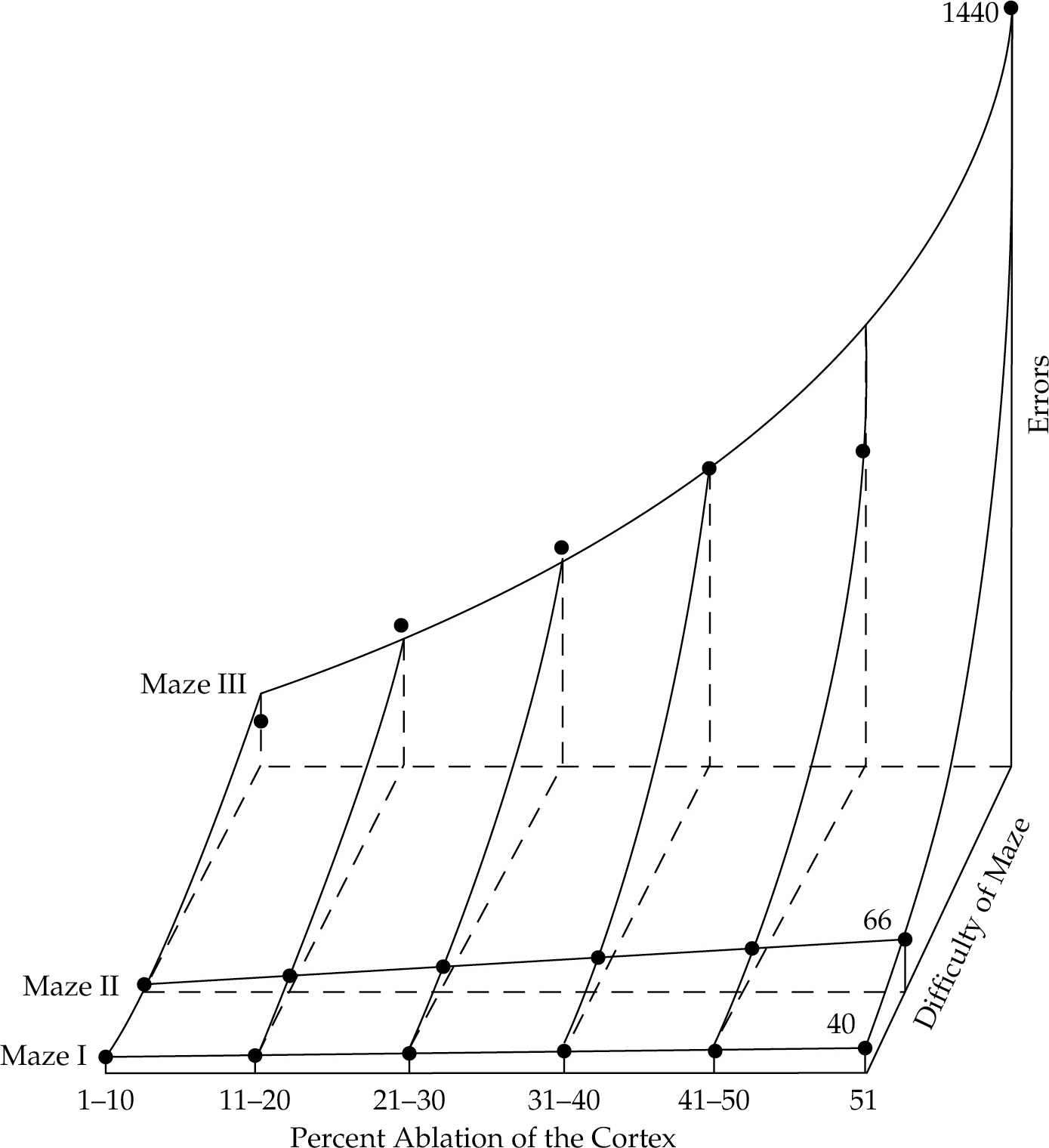
Figure 3.9 The results of Lashley’s ablation studies.
Just as Broca, Ferrier, and Wernicke had revived the localizationist argument, Lashley now harkened back to Flourens’s notion of the brain’s common action (action commune), although for Lashley it was memory rather than the will that seemed evenly distributed throughout the cortex. In summarizing his results, Lashley contributed two new terms of which Flourens would undoubtedly have approved. He said the brain is marked by equipotentiality, which he defined as “the apparent capacity of any intact part of a functional brain to carry out . . . the [memory] functions which are lost by the destruction of [other parts].” In other words, the brain has sufficient neural plasticity so that when one part of it is injured, other parts can potentially take over in providing the same functions. Sometimes, however, the brain’s equipotentiality may be offset by the law of mass action, “whereby the efficiency of performance of an entire complex function may be reduced in proportion to the extent of brain injury.”14 Simply put: the more extensive the brain injury, the less the opportunity for equipotentiality to operate.
In sum, Lashley’s systematic experiments seemed to rule out any simple theory of memory localization in the brain, and toward the end of his life he wryly expressed his own frustration with the problem: “I sometimes feel in reviewing the evidence on the localization of [memory]. . . that the necessary conclusion is that learning just is not possible. It is difficult to conceive of a mechanism which can satisfy the conditions set for it.”15
The exact mechanisms of memory remain something of a mystery today. Some of Lashley’s successors have suggested that his original theory oversimplified the problem, and that even elementary maze learning actually involves much more than the coupling of single sensory stimuli with single motor responses. In learning to run a maze, a rat must inevitably associate many different stimuli (involving touch, smell, and hearing as well as vision) with the various “correct” motor responses. Even if localized stimulus-response connections really do underlie learning and memory, there should be many of them in many different parts of the brain for any single completed act of maze learning. Damage to just a small part of the brain would remove just a few of these and would have a small effect on overall learning; damage to larger areas would remove a larger portion of the total, and produce a larger decrease in performance.
The so-called redundancy hypothesis offers a related explanation, suggesting that each individual memory gets stored in several locations throughout the cortex, with the number increasing as the memory becomes better established and more widely associated with other memories. Ablation of an isolated brain area would be expected to remove some but not all of the traces of any particular memory.
Despite the plausibility of these ideas, a definitive theory linking memory to specific brain activities has yet to be established. When it happens, it will almost certainly rely on new techniques that were perfected in the decades following Lashley’s research involving not the ablation, but rather the electrical stimulation of the conscious human brain.