QUESTION 2–6
Why do you suppose only ʟ-amino acids, and not a random mixture of the ʟ- and ᴅ-forms of each amino acid, are used to make proteins?
Having looked at the ways atoms combine to form small molecules and how these molecules behave in an aqueous environment, we now examine the main classes of small molecules found in cells and their biological roles. Amazingly, we will see that a few basic categories of molecules, formed from just a handful of different elements, give rise to all the extraordinary richness of form and behavior displayed by living things.
If we disregard water, nearly all the molecules in a cell are based on carbon. Carbon is outstanding among all the elements in its ability to form large molecules. Because a carbon atom is small and has four electrons and four vacancies in its outer shell, it readily forms four covalent bonds with other atoms (see Figure 2–9). Most importantly, one carbon atom can link to other carbon atoms through highly stable covalent C–C bonds, producing rings and chains that can form the backbone of complex molecules with no obvious upper limit to their size. These carbon-containing compounds are called organic molecules. By contrast, all other molecules, including water, are said to be inorganic.
In addition to containing carbon, the organic molecules produced by cells frequently contain specific combinations of atoms, such as the methyl (–CH3), hydroxyl (–OH), carboxyl (–COOH), carbonyl (–C=O), phosphoryl (–PO32–), and amino (–NH2) groups. Each of these chemical groups has distinct chemical and physical properties that influence the behavior of the molecule in which the group occurs, including whether the molecule tends to gain or lose protons when dissolved in water and with which other molecules it will interact. Knowing these groups and their chemical properties greatly simplifies understanding the chemistry of life. The most common chemical groups and some of their properties are summarized in Panel 2–1 (pp. 70–71).
The small organic molecules of the cell are compounds with masses in the range of 100–1000 daltons that contain up to 30 or so carbon atoms. They are usually found free in solution and have many different roles. Some are used as monomer subunits to construct the cell’s polymeric macromolecules—its proteins, nucleic acids, and large polysaccharides. Others serve as energy sources, being broken down and transformed into other small molecules in a maze of intracellular metabolic pathways. Many have more than one role in the cell—acting, for example, as both a potential subunit for a macromolecule and as an energy source. The small organic molecules are much less abundant than the organic macromolecules, accounting for only about one-tenth of the total mass of organic matter in a cell. But small organic molecules adopt a huge variety of chemical forms. Nearly 4000 different kinds of small organic molecules have been detected in the well-studied bacterium Escherichia coli.
All organic molecules are synthesized from—and are broken down into—the same set of simple compounds. Both their synthesis and their breakdown occur through sequences of chemical changes that are limited in variety and follow step-by-step rules. As a consequence, the compounds in a cell are chemically related, and most can be classified into a small number of distinct families. Broadly speaking, cells contain four major families of small organic molecules: the sugars, the fatty acids, the amino acids, and the nucleotides (Figure 2–17). Although many compounds present in cells do not fit into these categories, these four families of small organic molecules—together with the macromolecules made by linking them into long chains—account for a large fraction of a cell’s mass (Table 2–2).
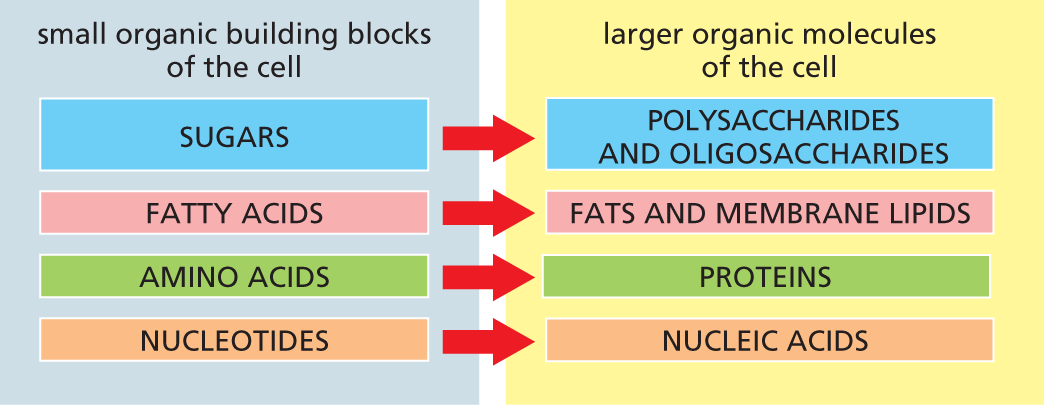
A chart shows the four major families of small organic molecules and their corresponding large organic molecules of the cell. The four major small organic building blocks of the cell are sugars, fatty acids, amino acids, and nucleotides. The large organic molecules of the cell are polysaccharides, oligosaccharides, fats, membrane lipids, proteins, and nucleic acids. Sugars are the subunits of polysaccharides and oligosaccharides; fatty acids are the subunits of fats and membrane lipids; amino acids are the subunits of proteins; nucleotides are the subunits of nucleic acids.
Figure 2–17 Sugars, fatty acids, amino acids, and nucleotides are the four main families of small organic molecules in cells. They form the monomeric building blocks, or subunits, for larger organic molecules, including the macromolecules and other molecular assemblies of the cell. Some, like the sugars and the fatty acids, are also energy sources. Oligosaccharides typically contain a smaller number of sugar subunits (disaccharides, for example, have only two), whereas polysaccharides such as glycogen (in animal cells) and starch (in plants) can contain thousands of sugar monomers.
|
TABLE 2–2 THE CHEMICAL COMPOSITION OF A BACTERIAL CELL |
||
|
Substance |
Percent of Total Cell Weight |
Approximate Number of Types in Each Class |
|
Water |
70 |
1 |
|
Inorganic ions |
1 |
20 |
|
Sugars and precursors |
1 |
250 |
|
Amino acids and precursors |
0.4 |
100 |
|
Nucleotides and precursors |
0.4 |
100 |
|
Fatty acids and precursors |
1 |
50 |
|
Other small molecules |
0.2 |
3000 |
|
Phospholipids |
2 |
4* |
|
Macromolecules (nucleic acids, proteins, and polysaccharides) |
24 |
3000 |
The simplest sugars—the monosaccharides—are compounds with the general formula (CH2O)n, where n is usually 3, 4, 5, or 6. Glucose, for example, has the formula C6H12O6 (Figure 2–18). Because of this simple formula, sugars, and the larger molecules made from them, are called carbohydrates. The formula, however, does not adequately define the molecule: the same set of carbons, hydrogens, and oxygens can be joined together by covalent bonds in a variety of ways, creating structures with different shapes. Thus glucose can be converted into a different sugar—mannose or galactose—simply by switching the orientations of specific –OH groups relative to the rest of the molecule. In addition, each of these sugars can exist in either of two forms, called the ᴅ-form and the ʟ-form, which are mirror images of each other. Sets of molecules with the same chemical formula but different structures are called isomers, and mirror-image pairs of such molecules are called optical isomers. Isomers are widespread among organic molecules in general, and they play a major part in generating the enormous variety of sugars. An outline of sugar structures and chemistry is presented in Panel 2–4 (pp. 76–77).
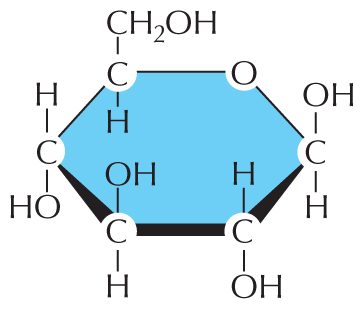
The first illustration is a Haworth projection of glucose. The glucose molecule is a hexagonal ring with a carbon atom, designated C, at each point of the ring. C 6 is substituted with an oxygen atom, designated O. At the first point to the right of O, is C 1 with a single bond to a hydroxyl group, O H, above the ring and a single bond to a hydrogen atom, H, below the ring. C 2 has a single bond to an H above the ring and a single bond to an O H group below the ring. C 3 has a single bond to an O H group above the ring and a single bond to an H group below the ring. C 4 has a single bond to an H above the ring and a single bond to an O H below the ring. C 5 has a single bond to H below the ring and a single bond to a C H subscript 2 O H group above the ring.
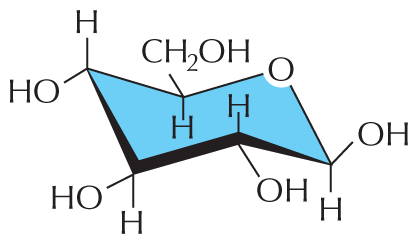
The second illustration is a chair configuration of glucose. The structure resembles a reclining beach chair with six points. One of the points in the structure has an O atom. At the first point to the right of the O atom, an O H group is shown branching above the chair and an H atom is shown branching below the chair. This point is the lowest in the chair configuration. The second point contains a bond to an H atom above the chair and to an O H group below the chair. The third point contains a bond to an O H group above the chair and to an H atom below the chair. The fourth point from the O atom has a bond to an H atom above the chair and to an O H group below the chair. This is the highest point in the structure. The fifth point has a bond to a C H subscript 2 O H group above the chair and to an H atom below the chair.
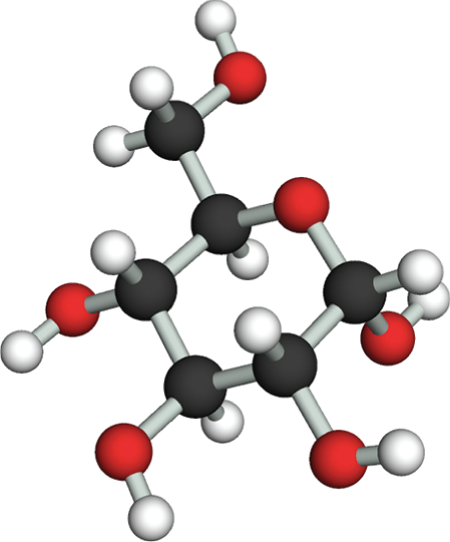
The third illustration is a ball and stick model of glucose. The model shows a central hexagonal ring with five carbon atoms, represented by black spheres, and one oxygen atom, represented by a red sphere. The first four carbon atoms to the right of the oxygen atom are each bound to a hydrogen atom, represented by a white sphere, and to a hydroxyl group, represented by a red sphere connected to a white sphere. At C 1 the H atom and the O H group are branching away from the carbon. At C 2 the H atom is above the carbon and the O H group is below the carbon. At C 3 the O H group is above the carbon and the H atom is below the carbon. At C 4 the H atom is above the carbon and the O H group is below the carbon. The fifth carbon is bound to a hydrogen below and a carbon above. The carbon above is bound to two hydrogen atoms and an O H group.
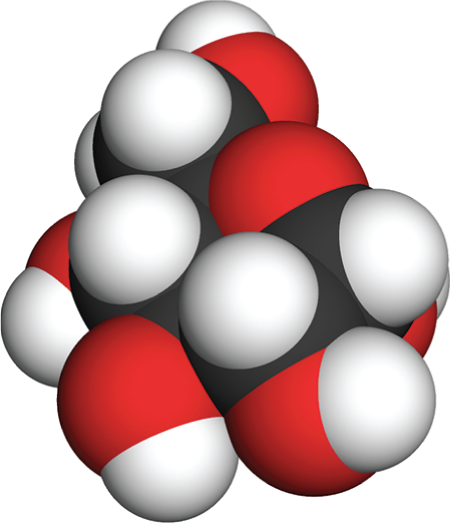
The fourth illustration is a space filling model of glucose. This model is composed of tightly packed spheres that follow conventional color-coding for the atoms of oxygen, carbon, and hydrogen. The packing of the spheres forms an irregular three dimensional shape. The hexagonal ring cannot be identified in this model. Incomplete spheres indicate the points at which the atoms are bound to each other. Spheres of hydrogen are shown to be smaller than spheres of oxygen or carbon.
Monosaccharides can be linked by covalent bonds—called glycosidic bonds—to form larger carbohydrates. Two monosaccharides linked together make a disaccharide, such as sucrose, which is composed of a glucose and a fructose unit. Larger sugar polymers range from the oligosaccharides, which typically contain 2 to 10 monosaccharide subunits (monomers), all the way up to giant polysaccharides, which can contain hundreds or thousands of subunits.
The way sugars are linked together illustrates some common features of biochemical bond formation. A bond is formed between an –OH group on one sugar and an –OH group on another by a condensation reaction, in which a molecule of water is expelled as the bond is formed (Figure 2–19). The subunits in other biological polymers, including nucleic acids and proteins, are also linked by condensation reactions in which water is expelled. The bonds created by these condensation reactions can subsequently be broken by the reverse process of hydrolysis, in which a molecule of water is consumed. Generally speaking, condensation reactions, which synthesize larger molecules from smaller subunits, are energetically unfavorable; hydrolysis reactions, which break down larger molecules into smaller subunits, are energetically favorable (Figure 2–20).
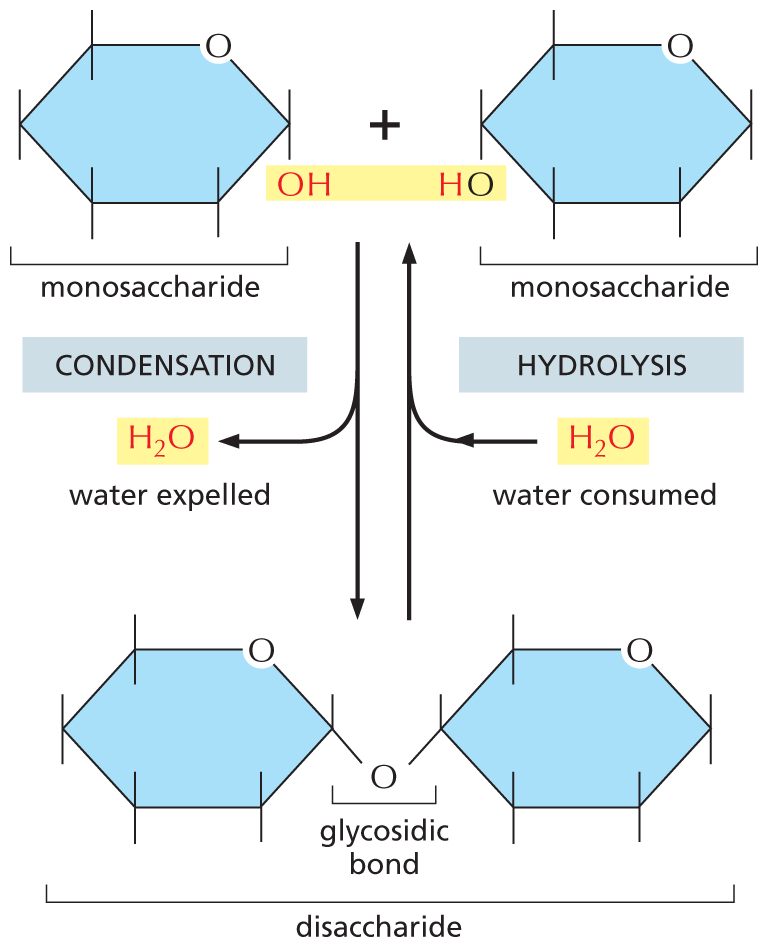
An illustration shows that a condensation reaction is the reverse of a hydrolysis reaction. There are two monosaccharides shown at the top of the figure. In the first monosaccharide, C 6 is replaced by an oxygen atom. C 1 is single bonded to an O H group (highlighted) below the ring and has a free single bond above the ring. C 2, C 3, C 4, and C 5 have each a free single bond above and below the ring. In the second monosaccharide, C 6 is replaced by an oxygen atom. C 4 is single bonded to an O H group below the ring in which H is highlighted and has a free single bond above the ring. C 1, C 2, C 3, and C 5 have each a free single bond above and below the ring. At the bottom of the figure is a disaccharide. Two six membered rings are shown, in which C 6 is replaced by an oxygen atom; C 1 of the first ring and C 4 of the second ring are each single bonded to an oxygen atom below the ring and the bond is labeled as a glycosidic bond. It is shown that the two monosaccharides form a disaccharide through a condensation reaction, in which water is expelled. The disaccharide with water can form the monosaccharides through a hydrolysis reaction, in which water is consumed.
Figure 2–19 Two monosaccharides can be linked by a covalent glycosidic bond to form a disaccharide. This reaction belongs to a general category of reactions termed condensation reactions, in which two molecules join together as a result of the loss of a water molecule. The reverse reaction (in which water is added) is termed hydrolysis.
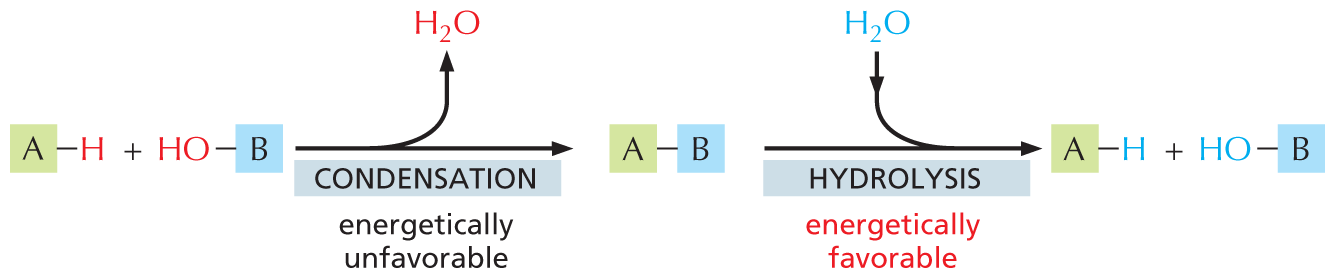
An illustration shows that condensation is energetically unfavorable and that hydrolysis is energetically favorable. A macromolecule labeled A with a single bond to a hydrogen atom undergoes energetically unfavorable condensation with a hydroxyl group single bonded to a macromolecule labeled B. A water molecule is released in the process. A macromolecule is formed in which A has a single bond to B.
This macromolecule undergoes energetically favorable hydrolysis through the addition of water to form macromolecule A with a single bond to a hydrogen atom and a hydroxyl group with a single bond to macromolecule B. It is indicated that in these reactions the hydrogen atom and the hydroxyl group are part of the H subscript 2 O molecule that is released during condensation and gained during hydrolysis.
Figure 2–20 Condensation and hydrolysis are reverse reactions. The large polymeric macromolecules of the cell are formed from subunits (or monomers) by condensation reactions, and they are broken down by hydrolysis. Condensation reactions are energetically unfavorable; thus macromolecule formation requires an input of energy, as we discuss in Chapter 3.
The monosaccharide glucose has a central role as an energy source for cells, as we explain in Chapter 13. It is broken down to smaller molecules in a series of reactions, releasing energy that the cell can harness to do useful work. Cells use simple polysaccharides composed only of glucose units—principally glycogen in animals and starch in plants—as long-term stores of glucose, held in reserve for energy production.
Sugars do not function exclusively in the production and storage of energy. They are also used, for example, to make mechanical supports. The most abundant organic molecule on Earth—the cellulose that forms plant cell walls—is a polysaccharide of glucose. Another extraordinarily abundant organic substance, the chitin of insect exoskeletons and fungal cell walls, is also a polysaccharide—in this case, a linear polymer of a sugar derivative called N-acetylglucosamine. Other polysaccharides, which tend to be slippery when wet, are the main components of slime, mucus, and gristle. The number of possible polysaccharides is vast, in part because monosaccharides have several free –OH groups that can participate in the formation of covalent bonds (see Figure 2–18A); thus, even sugar polymers made from the same monosaccharides can form a variety of complex, branched structures (see Panel 2–4).
Smaller oligosaccharides can be covalently linked to proteins to form glycoproteins, or to lipids to form glycolipids, both of which can be found in cell membranes. The sugar side chains attached to glycoproteins and glycolipids in the plasma membrane are thought to help protect the cell surface and often help cells adhere to one another. Differences in the types of cell-surface sugars form the molecular basis for the human blood groups (see Panel 2–4); this information dictates which blood types can be used during transfusions.
A fatty acid molecule, such as palmitic acid, has two chemically distinct regions. One is a long hydrocarbon chain, which is hydrophobic and not very reactive chemically. The other is a carboxyl (–COOH) group, which behaves as an acid (carboxylic acid): in an aqueous solution, it is ionized (–COO–), extremely hydrophilic, and chemically reactive (Figure 2–21). Molecules—such as fatty acids—that possess both hydrophobic and hydrophilic regions are termed amphipathic. Almost all the fatty acid molecules in a cell are covalently linked to other molecules by their carboxylic acid group. Fatty acids and other lipids are reviewed in Panel 2–5 (pp. 78–79).
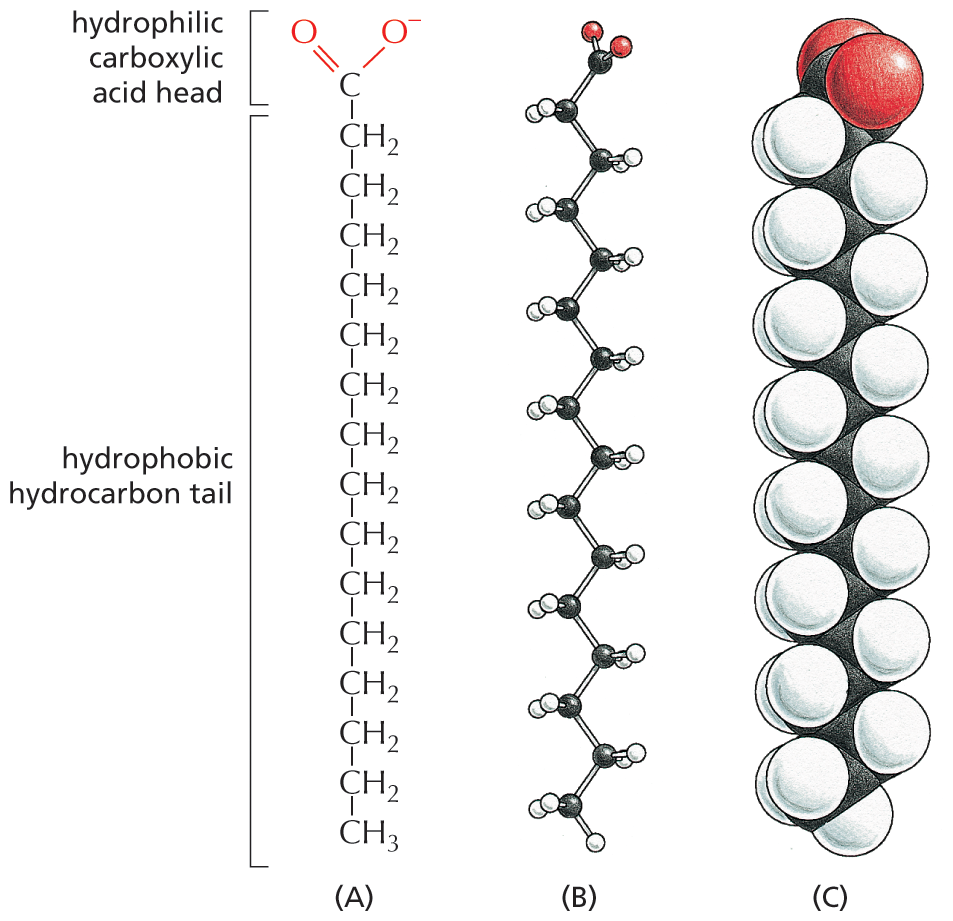
Three illustrations show the chemical structure, ball and stick model, and space filling model of a fatty acid. Each illustration is arranged with the hydrophilic carboxylic acid head of the fatty acid at the top and the hydrophobic hydrocarbon tail at the bottom.
Figure 2–21 Fatty acids have both hydrophobic and hydrophilic components. The hydrophobic hydrocarbon chain is attached to a hydrophilic carboxylic acid group. Different fatty acids have different hydrocarbon tails. Palmitic acid is shown here. (A) Structural formula, showing the carboxylic acid head group in its ionized form, as it exists in water at pH 7. (B) Ball-and-stick model. (C) Space-filling model (Movie 2.2).
The hydrocarbon tail of palmitic acid is saturated: it contains the maximum possible number of hydrogens and therefore has no double bonds between its carbon atoms. Some other fatty acids, such as oleic acid, have unsaturated tails, with one or more double bonds along their length. The double bonds create kinks in the hydrocarbon tails, which interferes with their ability to pack together. When these fatty acid tails are located in a cell membrane, the tightness of their packing affects the membrane’s fluidity. The many different fatty acids found in cells differ only in the length of their hydrocarbon chains and in the number and position of the carbon–carbon double bonds (see Panel 2–5).
Fatty acids serve as a concentrated food reserve in cells: they can be broken down to produce about six times as much usable energy, gram for gram, as glucose. Fatty acids are stored in the cytosol of many cells in the form of fat droplets composed of triacylglycerol molecules—compounds made of three fatty acid chains covalently joined to a glycerol molecule (Figure 2–22; see also Panel 2–5). Triacylglycerols are the animal fats found in meat, butter, and cream, and the plant oils such as corn oil and olive oil. When a cell needs energy, the fatty acid chains can be released from triacylglycerols and broken down into two-carbon units. These two-carbon units are identical to those derived from the breakdown of glucose, and they enter the same energy-yielding reaction pathways, as described in Chapter 13.
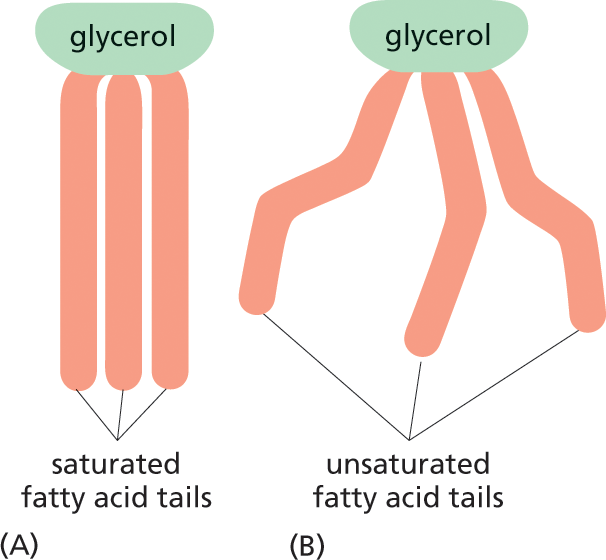
Two illustrations show the structures of saturated and unsaturated fatty acid tails of glycerol.
Figure 2–22 The properties of fats depend on the length and saturation of the fatty acid chains they carry. Fatty acids are stored in the cytosol of many cells in the form of droplets of triacylglycerol molecules made of three fatty acid chains joined to a glycerol molecule. (A) Saturated fats are found in meat and dairy products. (B) Plant oils, such as corn oil, contain unsaturated fatty acids, which may be monounsaturated (containing one double bond) or polyunsaturated (containing multiple double bonds). The presence of these double bonds causes plant oils to be liquid at room temperature. Although fats are essential in the diet, saturated fats raise the concentration of cholesterol in the blood, which tends to clog the arteries, increasing the risk of heart attacks and strokes.
Fatty acids and their derivatives, including triacylglycerols, are examples of lipids. Lipids are loosely defined as molecules that are insoluble in water but soluble in fat and organic solvents such as benzene. They typically contain long hydrocarbon chains, as in the fatty acids, or multiple linked aromatic rings, as in the steroids (see Panel 2–5).
Among the most important lipids in the cell are the phospholipids. Like triacylglycerols, most phospholipids are constructed mainly from fatty acids and glycerol. In phospholipids, however, the glycerol is joined to two fatty acid chains, rather than to three as in triacylglycerols. The remaining –OH group on the glycerol is linked to a hydrophilic phosphate group, which in turn is attached to a small hydrophilic compound such as choline (Figure 2–23A).
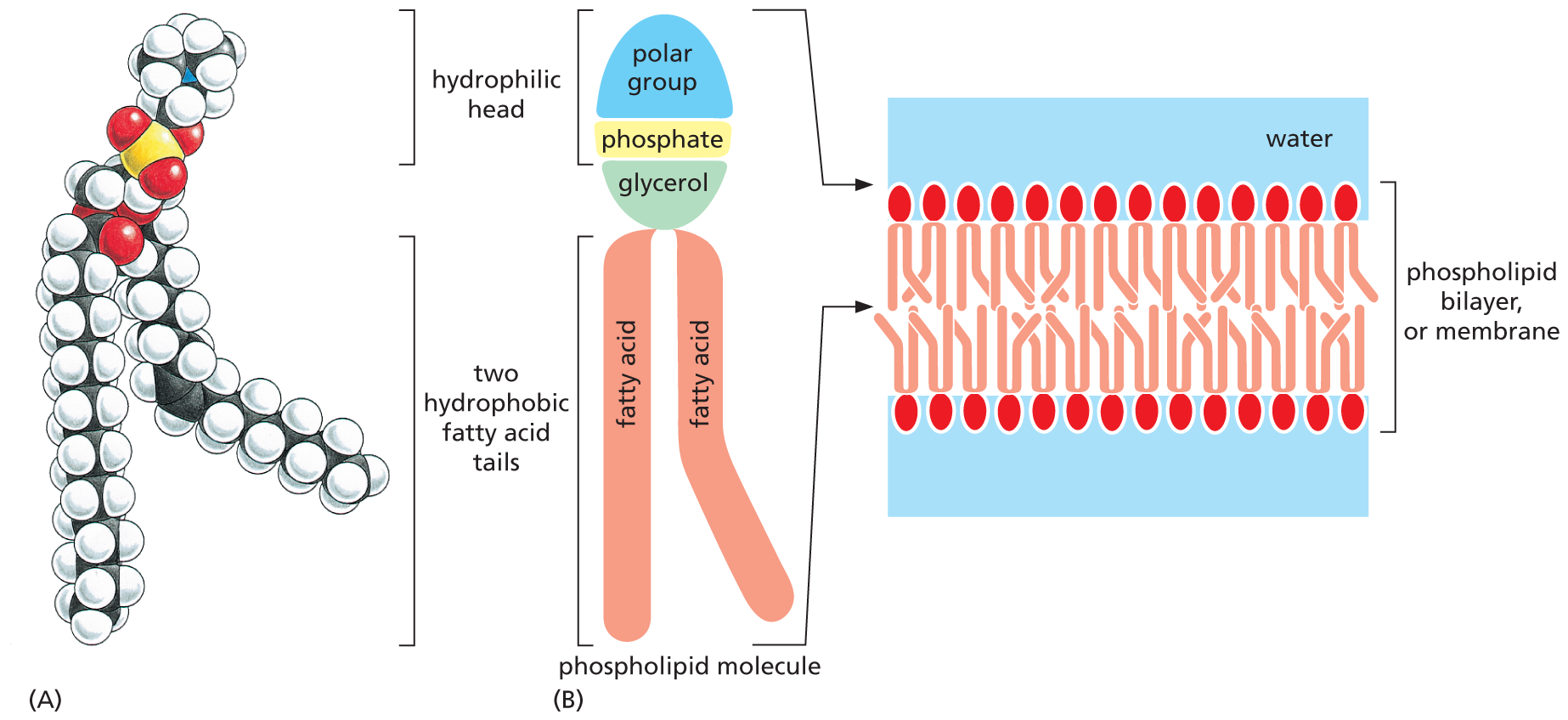
Two illustrations show the structure of the phospholipid phosphatidylcholine and the membrane arrangement of phospholipids.
Figure 2–23 Phospholipids can aggregate to form cell membranes. Phospholipids contain two hydrophobic fatty acid tails and a hydrophilic head. (A) Phosphatidylcholine is the most common phospholipid in cell membranes. The phosphorus atom is shown in yellow and oxygen atoms in red. (B) Diagram showing how, in an aqueous environment, the hydrophobic tails of phospholipids pack together to form a lipid bilayer. In the lipid bilayer, the hydrophilic heads of the phospholipid molecules are on the outside, facing the aqueous environment, and the hydrophobic tails are on the inside, where water is excluded.
With their hydrophilic, phosphate-containing head and hydrophobic fatty acid tails, phospholipids are strongly amphipathic. This characteristic composition and shape gives them very different physical and chemical properties from triacylglycerols, which are predominantly hydrophobic. It is thanks to their amphipathic nature that pure phospholipids readily form membranes in water. They can spread over the surface of water to form a monolayer, with their hydrophobic tails facing the air and their hydrophilic heads in contact with the water. Alternatively, two phospholipid layers can combine tail-to-tail in water to form a phospholipid sandwich called a lipid bilayer (Figure 2–23B). This structure forms the basis for the membranes that surround every cell and enclose their internal organelles. Although they are composed largely of phospholipids, cell membranes also contain differing amounts of other lipids, including glycolipids, which are structurally similar to phospholipids but contain one or more sugars instead of a phosphate group.
Amino acids are small organic molecules with one defining property: they all possess a carboxylic acid group (–COOH) and an amino group (–NH2), both attached to a central α-carbon atom (Figure 2–24). This α-carbon also carries a specific side chain, the identity of which distinguishes one amino acid from another.
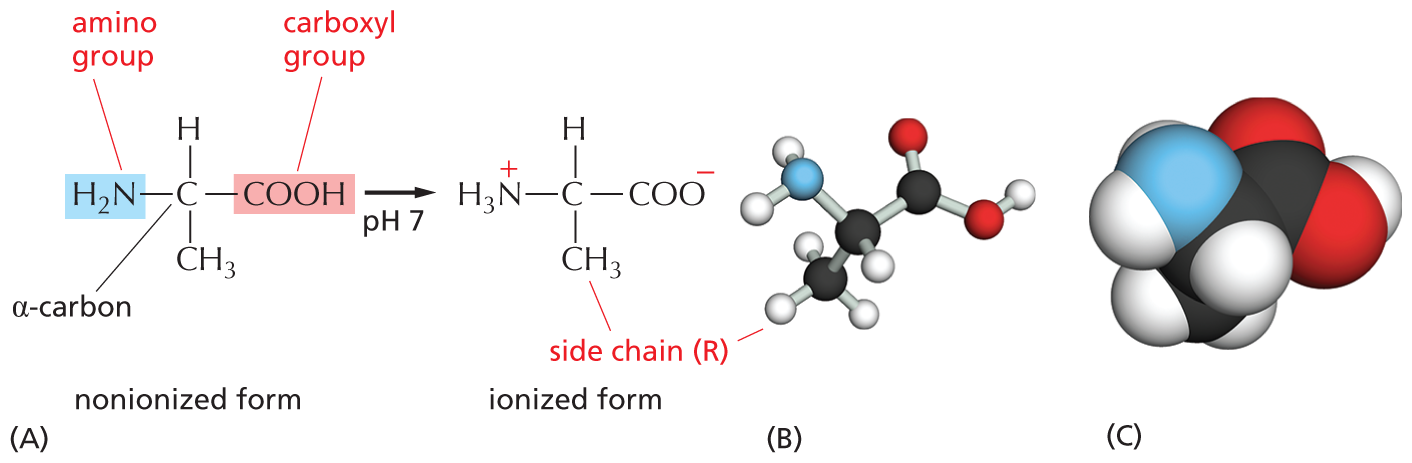
Three illustrations show ionization of alanine, a ball and stick model of alanine, and a space filling model of alanine.
Figure 2–24 All amino acids have an amino group, a carboxyl group, and a side chain (R) attached to their α-carbon atom. In the cell, where the pH is close to 7, free amino acids exist in their ionized form; but, when they are incorporated into a polypeptide chain, the charges on their –NH2 and –COOH groups are lost. (A) The amino acid shown is alanine, one of the simplest amino acids, which has a methyl group (–CH3) as its side chain. Its amino group is highlighted in blue and its carboxyl group in red. (B) A ball-and-stick model and (C) a space-filling model of alanine. In (B) and (C), the N atom is blue and the O atom is red.
Cells use amino acids to build proteins—polymers made of amino acids, which are joined head-to-tail in a long chain that folds up into a three-dimensional structure that is unique to each type of protein. The covalent bond between two adjacent amino acids in a protein chain is called a peptide bond, and the resulting chain of amino acids is therefore also known as a polypeptide. Peptide bonds are formed by condensation reactions that link one amino acid to the next. Regardless of the specific amino acids from which it is made, the polypeptide always has an amino group at one end—its N-terminus—and a carboxyl group at its other end—its C-terminus (Figure 2–25). This difference in the two ends gives a polypeptide a definite directionality—a structural (as opposed to electrical) polarity.
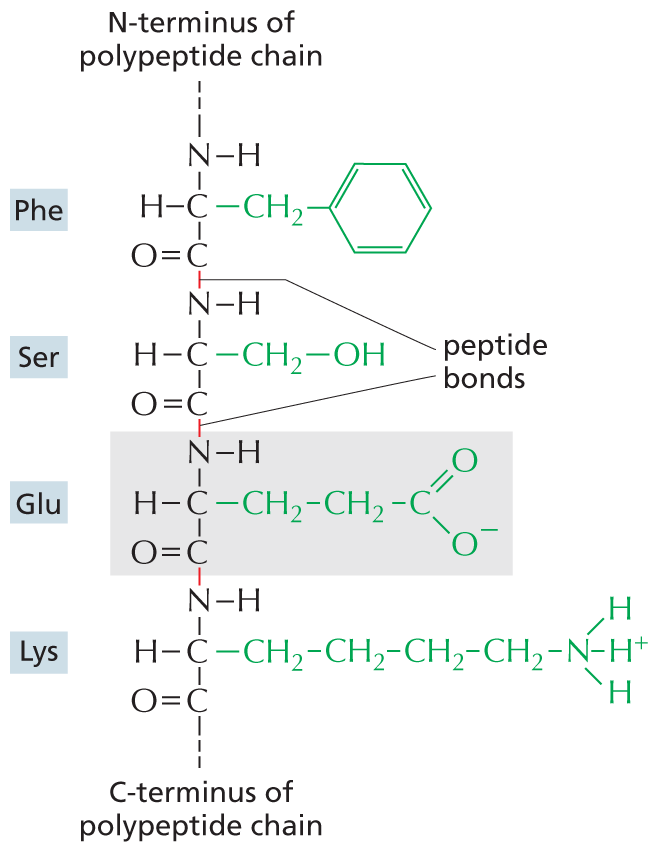
An amino acid chain is shown, with an emphasis on the peptide bonds and side chains. From top to bottom, the amino acids in the chain are phenylalanine, serine, glutamic acid, and lysine. In phenylalanine, shortened to P H E, two carbon atoms are single bonded to each other. One of the carbon atoms is single bonded to hydrogen, an N H group, and C H subscript 2. The C H subscript 2 group with a single bond to a benzene ring makes up the amino acid side chain in phenylalanine. The other carbon atom is double bonded to an oxygen atom and single bonded to the N H group of serine. The bond between the carbon atom of phenylalanine and the nitrogen atom of serine is identified as a peptide bond.
In serine, shortened to S E R, two carbon atoms are single bonded to each other. The first carbon atom is single bonded to a hydrogen atom, an N H group, and C H subscript 2. The C H subscript 2 group with a single bond to an O H group makes up the amino acid side chain in serine. The other carbon atom is double bonded to an oxygen atom and to the N H group of glutamic acid. The bond between the carbon atom of serine and the nitrogen atom of glutamic acid is identified as a peptide bond.
In glutamic acid, shortened to G L U, two carbon atoms are single bonded to each other. The first carbon atom is single bonded to a hydrogen atom, an N H group, and C H subscript 2. The amino acid side chain in glutamic acid consists of this C H subscript 2 group single bonded to a C H subscript 2 group that has a single bond to a carbon atom. The side chain carbon atom is double bonded to oxygen and single bonded to an oxygen with a negative charge. The other carbon atom is double bonded to oxygen and to the N H group of lysine. The bond between the carbon atom of glutamic acid and the nitrogen atom of lysine is identified as a peptide bond.
In lysine, shortened to L Y S, two carbon atoms are single bonded to each other. The first carbon is single bonded to an N H group, a hydrogen atom, and C H subscript 2. The amino acid side chain in lysine consists of the C H subscript 2 group single bonded to a chain of three C H subscript 2 groups. The terminal C H subscript 2 group has a single bond to nitrogen. The side chain nitrogen atom has three single bonds to three hydrogen atoms in which the central hydrogen atom is positively charged. The other carbon atom of lysine is double bonded to an oxygen atom. The nitrogen of P H E is single bonded to the N-terminus of the polypeptide chain and the carbon of L Y S is single bonded to the C-terminus of the polypeptide chain.
Figure 2–25 Amino acids in a protein are held together by peptide bonds. The four amino acids shown are linked together by peptide bonds (red). One of the amino acids, glutamic acid, is shaded in gray. The amino acid side chains are shown in green. The N-terminus of the polypeptide chain is capped by an amino group, and the C-terminus ends in a carboxyl group. The sequence of amino acids in a protein is abbreviated using either a three-letter or a one-letter code, and the sequence is always read starting from the N-terminus (see Panel 2–6, pp. 80–81). In the example given, the sequence is Phe-Ser-Glu-Lys (or FSEK).
Twenty types of amino acids are commonly found in proteins, each with a different side chain attached to its α-carbon atom. The structures of all of them are displayed in Panel 2–6 (pp. 80–81). How this precise set of 20 amino acids came to be chosen is one of the mysteries surrounding the evolution of life; there is no obvious chemical reason why other amino acids could not have served just as well. But once the selection had been locked into place, it could not be changed, as too much chemistry had evolved to exploit it.
Like sugars, all amino acids (except glycine) exist as optical isomers termed ᴅ- and ʟ-forms (see Panel 2–6). But only ʟ-forms are ever found in proteins (although ᴅ-amino acids occur as part of bacterial cell walls and in some antibiotics, and ᴅ-serine is used as a signal molecule in the brain). The origin of this exclusive use of ʟ-amino acids to make proteins is another evolutionary mystery.
QUESTION 2–6
Why do you suppose only ʟ-amino acids, and not a random mixture of the ʟ- and ᴅ-forms of each amino acid, are used to make proteins?
The chemical versatility that the 20 standard amino acids provide is vitally important to the function of proteins. Five of the 20 amino acids—including lysine and glutamic acid, shown in Figure 2–25—have side chains that form ions in solution and can therefore carry a charge. The others are uncharged. Some amino acids are polar and hydrophilic, and some are nonpolar and hydrophobic (see Panel 2–6). As we discuss in Chapter 4, the collective properties of the amino acid side chains underlie all the diverse and sophisticated functions of proteins. And proteins, which constitute half the dry mass of a cell, lie at the center of life’s chemistry.
DNA and RNA are built from subunits called nucleotides. Nucleotides consist of a nitrogen-containing ring linked to a five-carbon sugar that has a phosphate group attached to it. The sugar can be either ribose or deoxyribose. Nucleotides containing ribose are known as ribonucleotides, and those containing deoxyribose are known as deoxyribonucleotides. The structures of nucleotides and their derivatives are illustrated in Panel 2–7 (pp. 82–83).
The nitrogen-containing ring compounds of all these molecules are generally referred to as bases for historical reasons: under acidic conditions, they can each bind an H+ (proton) and thereby increase the concentration of OH– ions in aqueous solution. There is a strong family resemblance between the different nucleotide bases. Cytosine (C), thymine (T), and uracil (U) are called pyrimidines, because they all derive from a six-membered pyrimidine ring; guanine (G) and adenine (A) are purines, which bear a second, five-membered ring fused to the six-membered ring. Each nucleotide is named after the base it contains (see Panel 2–7). A base plus its sugar (without any phosphate group attached) is called a nucleoside.
Nucleoside di- and triphosphates can act as short-term carriers of chemical energy. Above all others, the ribonucleoside triphosphate known as adenosine triphosphate, or ATP (Figure 2–26), participates in the transfer of energy in hundreds of metabolic reactions. ATP is formed through reactions that are driven by the energy released from the breakdown of foodstuffs. Its three phosphates are linked in series by two phosphoanhydride bonds (see Panel 2–7). Rupture of these phosphate bonds by hydrolysis releases large amounts of useful energy, also known as free energy (see Panel 3–1, pp. 100–101). Most often, it is the terminal phosphate group that is split off—or transferred to another molecule—to release energy that can be used to drive biosynthetic reactions (Figure 2–27). Other nucleotide derivatives serve as carriers for other chemical groups. These reactions are described in detail in Chapter 3.
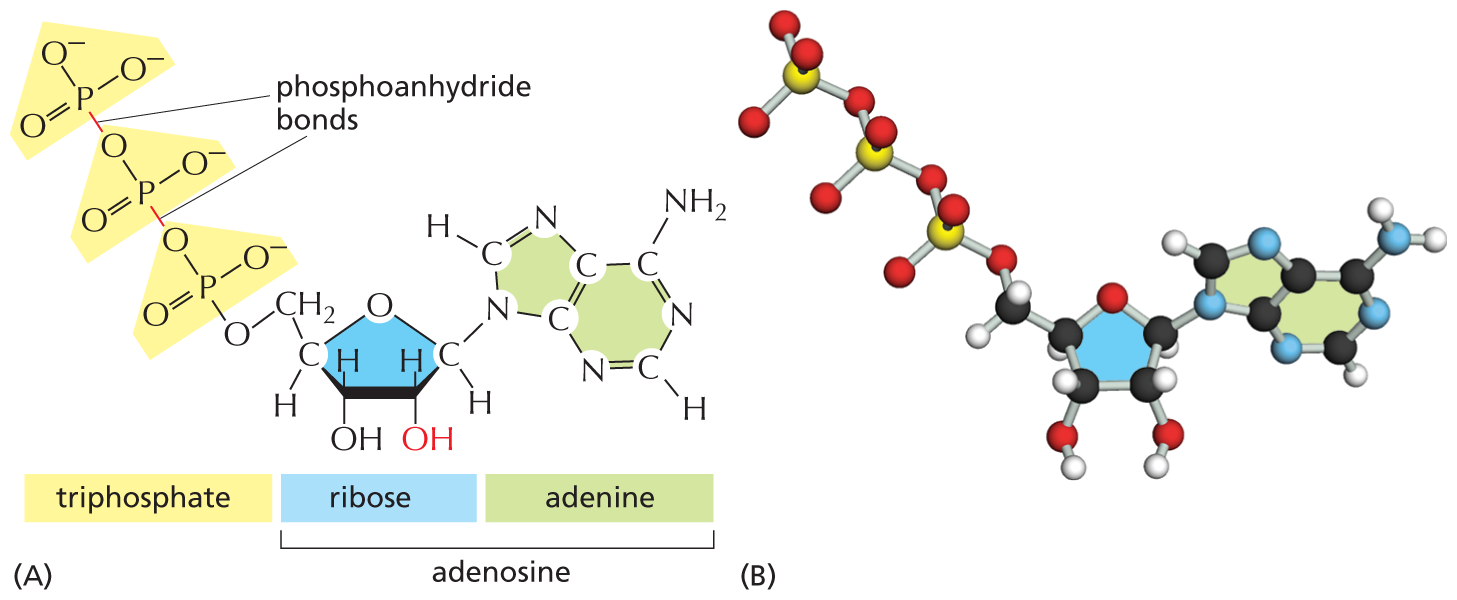
Two illustrations show the structure of A T P and its ball and stick model. Part A depicts the structural formula of Adenosine triphosphate. Part B depicts the ball and stick model of Adenosine triphosphate.
Figure 2–26 Adenosine triphosphate (ATP) is a crucially important energy carrier in cells. (A) Structural formula, in which the three phosphate groups are shaded in yellow. The presence of the –OH group on the second carbon of the sugar ring (red) distinguishes this sugar as ribose. (B) Ball-and-stick model (Movie 2.3). In (B), the P atoms are yellow.
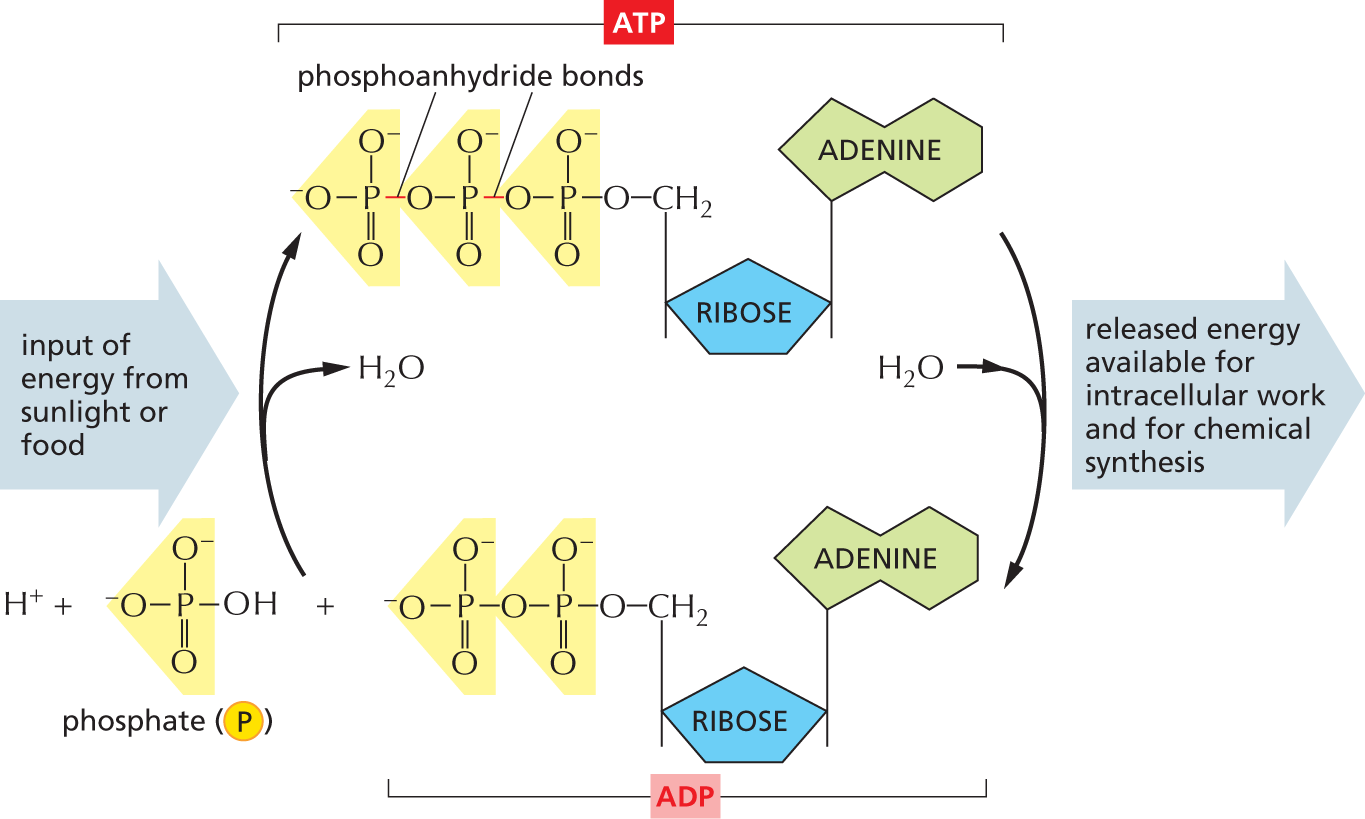
An illustration shows the synthesis and hydrolysis of A T P. A T P is synthesized through the reaction between a hydrogen ion, inorganic phosphate, and A D P. This reaction requires an input of energy from sunlight or food. A water molecule is released in the process of A T P synthesis. When A T P is hydrolyzed for energy release, A D P and phosphate remain. A water molecule is gained during the process of A T P hydrolysis. The energy released during A T P hydrolysis is available for intracellular work and for chemical synthesis.
The structures of the reactants and products are as follows:
The structure of inorganic phosphate shows a central phosphorous atom with single bonds to two oxygen ions that are negatively charged and a single bond to an O H group, and a double bond to an oxygen atom.
The structure of A D P shows an adenine single bonded to a ribose that is bonded to a diphosphate.
The structure of A T P shows a triphosphate linked by phosphoanhydride bonds bonded to a ribose unit that is further bonded to adenine.
Figure 2–27 ATP is synthesized from ADP and phosphate, and it releases energy when it is hydrolyzed back to ADP and phosphate. The energy required for ATP synthesis is derived from either the energy-yielding oxidation of foodstuffs (in animal cells, fungi, and some bacteria) or the capture of light (in plant cells and some bacteria). The hydrolysis of ATP releases energy that is used to drive many processes inside cells. Together, the two reactions shown form the ATP cycle.
Nucleotides also have a fundamental role in the storage and retrieval of biological information. They serve as building blocks for the construction of nucleic acids—long polymers in which nucleotide subunits are linked by the formation of covalent phosphodiester bonds between the phosphate group attached to the sugar of one nucleotide and a hydroxyl group on the sugar of the next nucleotide (Figure 2–28). Nucleic acid chains are synthesized from energy-rich nucleoside triphosphates by a condensation reaction that releases pyrophosphate—a pair of phosphate groups linked by a single phosphoanhydride bond (see Panel 2–7).
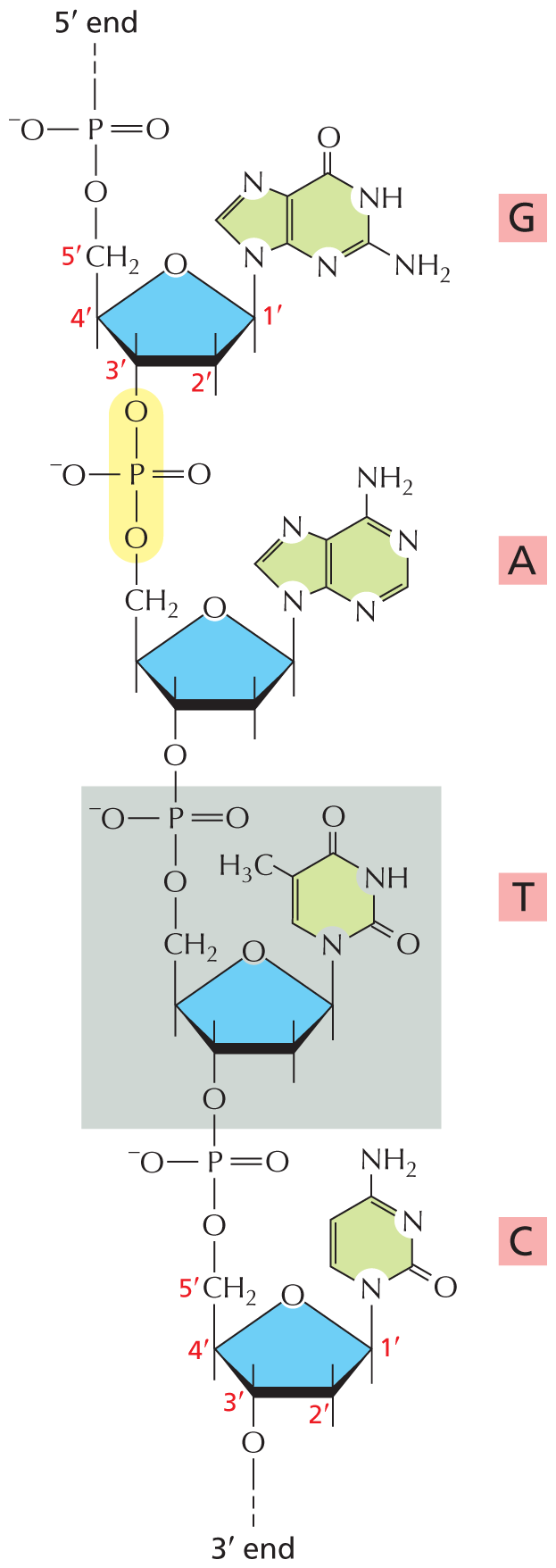
An illustration shows a short D N A molecule. In Guanine, a five membered ring is bonded to a six membered ring in which C 1 and C 2, C 5 and C 9, and C 6 and C 7 are double bonded. C 1, C 6, and C 8 are replaced by a nitrogen atom. C 2 is single bonded to N H subscript 2 group. C 3 is replaced by N H group. C 4 is double bonded to an oxygen atom. Another five membered ring in which C 5 is replaced by an oxygen atom is shown. C 1 is single bonded to C 8 of a five membered ring (highlighted in green color) above the ring and has a free single bond below the ring. C 2 has a free single bond above and below the ring. C 3 has a free single bond above the ring. C 4 has a free single bond below the ring and is single bonded to C H subscript 2 group above the ring. C 1, C 2, C 3, C 4, and C H subscript 2 are labeled as 1 prime, 2 prime, 3 prime, 4 prime, and 5 prime in red color. C H subscript 2 is single bonded to an oxygen atom which is single bonded to a phosphorus atom. The phosphorous atom is single bonded to an oxygen ion which is negatively charged, double bonded to an oxygen atom, and has a free bond which is labeled 5 prime end.
In Adenine, a five membered ring is bonded with a six membered ring in which C 1 and C 2, C 3 and C 4, C 5 and C 9, and C 6 and C 7 are double bonded to each other. C 1, C 3, C 6, and C 8 are replaced by a nitrogen atom. C 4 is single bonded to N H subscript 2 group. C 8 is single bonded to C 1 above the ring of a five membered ring. In the five membered ring, C 1 has a free single bond below the ring and C 2 has a free single bond above and below the ring. C 3 has a free single bond above the ring and is single bonded to an oxygen atom below the ring. C 4 has a free single bond below the ring and is single bonded to C H subscript 2 above the ring. C H subscript 2 is single bonded to an oxygen atom which is single bonded to a phosphorous atom. The phosphorous atom is single bonded to an oxygen ion which is negatively charged and an oxygen atom, double bonded to an oxygen atom. The phosphorous atom single bonded to the oxygen atoms is highlighted in yellow.
In Thymine, a six membered ring in which C 5 and C 6 are double bonded. C 1 is replaced by a nitrogen atom. C 2 and C 4 are doubled bonded to an oxygen atom. C 3 is replaced by N H group. C 5 is single bonded to C H subscript 3 group. C 1 is single bonded to C 1 above the ring of a five membered ring (highlighted in blue color). In the five membered ring, C 1 has a free single bond below the ring. C 3 has a free single bond above the ring and is single bonded to an oxygen atom below the ring. C 4 has a free single bond below the ring and is single bonded to C H subscript 2 above the ring. C H subscript 2 is single bonded to an oxygen atom which is single bonded to a phosphorous atom. The phosphorous atom is single bonded to an oxygen ion which is negatively charged and double bonded to an oxygen atom. The structure of thymine is highlighted in a grey.
Cytosine shows a six membered ring in which C 3 and C 4 and C 5 and C 6 are double bonded to each other. C 1 and C 3 are replaced by a nitrogen atom. C 2 is doubled bonded to an oxygen atom. C 4 is single bonded to N H subscript 2 group. In the five membered ring, C 1 has a free single bond below the ring. C 2 has a free single bond above and below the ring. C 3 has a free single bond above the ring. C 4 has a free single bond below the ring and is single bonded to C H subscript 2 above the ring. C 5 is replaced by an oxygen atom. C 1, C 2, C 3, C 4, and C H subscript 2 are labeled as 1 prime, 2 prime, 3 prime, 4 prime, and 5 prime. C H subscript 2 is single bonded to an oxygen atom which is single bonded to a phosphorous atom. The phosphorous atom is single bonded to an oxygen ion which is negatively charged and double bonded to an oxygen atom. The phosphorous atom is also single bonded to an oxygen atom that is single bonded to C 3 of a five membered ring of thymine.
Figure 2–28 A short length of one chain of a deoxyribonucleic acid (DNA) molecule shows the covalent phosphodiester bonds linking four consecutive nucleotides. Because these bonds involve specific carbon atoms in the sugar ring—known as the 5ʹ and 3ʹ carbon atoms—a polynucleotide chain is said to have a 5ʹ and 3ʹ end; the 5ʹ end has a free phosphate group and the 3ʹ end has a free hydroxyl group. One of the nucleotides, T, is shaded in gray, and one phosphodiester bond is highlighted in yellow. The linear sequence of nucleotides in a polynucleotide chain is commonly abbreviated using a one-letter code, and the sequence is always read from the 5ʹ end. In the example illustrated, the sequence is GATC.
There are two main types of nucleic acids, which differ in the type of sugar contained in their sugar–phosphate backbone. Those based on the sugar ribose are known as ribonucleic acids, or RNA, and contain the bases A, G, C, and U. Those based on deoxyribose (in which the hydroxyl group at the 2ʹ position of the ribose carbon ring is replaced by a hydrogen) are known as deoxyribonucleic acids, or DNA, and contain the bases A, G, C, and T (T is chemically similar to the U in RNA; see Panel 2–7). RNA usually occurs in cells in the form of a single-stranded polynucleotide chain that can fold up into a variety of shapes. DNA, in contrast, is virtually always double-stranded: the DNA double helix is composed of two polynucleotide chains that run in opposite directions and are held together by hydrogen bonds between the bases of the two chains (see Panel 2–3, pp. 74–75).
The linear sequence of nucleotides in a DNA or RNA molecule encodes genetic information. The two nucleic acids, however, have different roles in the cell. DNA, with its more stable, hydrogen-bonded double helix, acts as a long-term repository for hereditary information, while single-stranded RNA molecules usually serve as more transient carriers of molecular instructions. The ability of the bases in different nucleic acid molecules to recognize and pair with each other by hydrogen-bonding (called base-pairing)—G with C, and A with either T or U—underlies all of heredity and evolution, as explained in detail in Chapter 5.
