The bewildering diversity of microbial life forms presented nineteenth-century microbiologists with a seemingly impossible task of classification. So little was known about life under the lens that natural scientists despaired of ever learning how to distinguish microbial species. The famous classifier of species, Swedish botanist Carl von Linné (Carolus Linnaeus, 1707–1778), called the microbial world “chaos.” Linnaeus would have been astonished to see the intricate family tree of life revealed by Carl Woese in the twentieth century, consisting of three domains: Bacteria, Archaea, and Eukarya (Fig. 1.29A). Of these domains, Archaea was unknown in Linnaeus’s time, and the idea that archaea gave rise to eukaryotes—including humans—would have been impossible to imagine.
A
More information
A diagram of the three domains of life and a photograph of Carl Woese holding a space filling model.
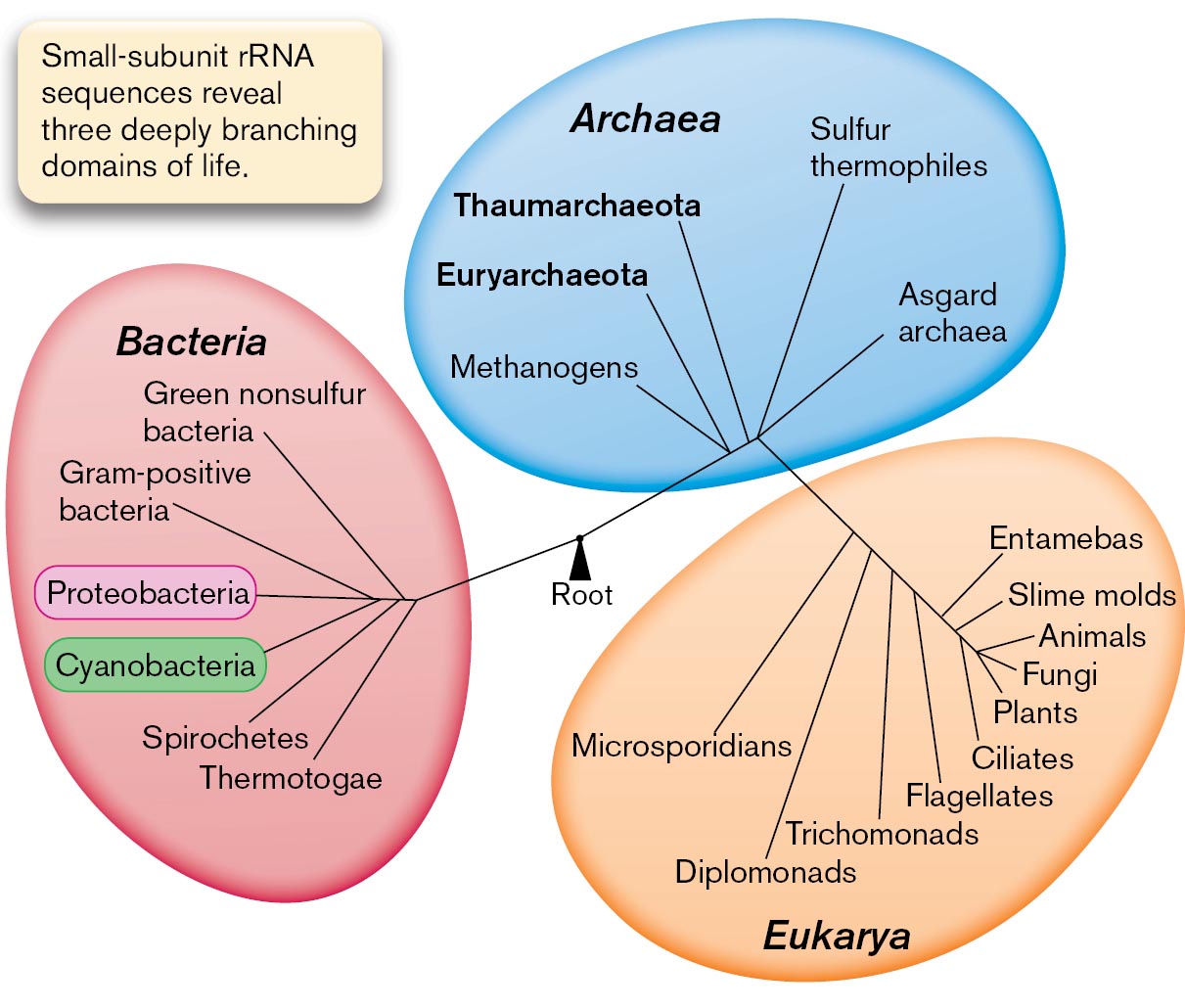
A diagram of the three domains of life: Archaea, Bacteria, and Eukarya. The three domains are connected by the root in the center, which is labeled as the last common ancestor. Text beside the diagram reads, Small-subunit r R N A sequences reveal three deeply branching domains of life. The first domain, Bacteria, is subdivided into several branches, namely, Green non-sulfur bacteria, Gram-positive bacteria, Proteobacteria, Cyanobacteria, Spirochetes, and Thermotogae. Archaea is subdivided into Methanogens, Euryarchaeota, Thaumarchaeota, Sulfur thermophiles, and Asgard archaea. The third domain shows Eukarya, which branches out into Microsporidians, Diplomonads, Trichomonads, Flagellates, Entamebas, Slime molds, Animals, Fungi, Plants, and Ciliates.
B
More information
A photo of Carl Woese holding a space filling model.
FIGURE 1.29 ■Carl Woese and the three domains of life.A. Three domains form a monophyletic tree that is based on small-subunit ribosomal RNA (rRNA) sequences. The length of each branch approximates the time of divergence from the last common ancestor. B. Carl Woese first used rRNA sequence to determine the phylogeny of living organisms.AP PHOTO
Microbes Are a Challenge to Classify
Before the twentieth century, taxonomists faced two challenges as they attempted to classify microbes. First, the resolution of the light microscope revealed little more than the outward shape of microbial cells, and vastly different kinds of microbes looked more or less alike (discussed in Chapter 2). This challenge was overcome as advances in biochemistry and microscopy made it possible to distinguish microbes by metabolism and cell structure, and ultimately by DNA sequence.
Second, microbes do not readily fit the classic definition of a species; that is, a group of organisms that interbreed. Unlike multicellular eukaryotes, microbes generally reproduce asexually. When they do exchange genes, they may do so with related strains or with distantly related species (discussed in Chapter 9). Nevertheless, microbiologists have devised working definitions of microbial species that enable us to usefully describe populations (discussed in Chapter 17). The most useful classification defines relatedness on the basis of similarity of DNA sequence: the more sequence two organisms share, the shorter the time since their populations diverged from a common ancestor. But early taxonomists had no DNA sequence information.
Note: The names of microbial species are frequently changed to reflect new understanding of genetic relationships. For example, the causative agent of bubonic plague was formerly called Bacterium pestis (1896), Bacillus pestis (1900), and Pasteurella pestis (1923), but it is now called Yersinia pestis (1944). The older names, however, still appear in the literature—a point to remember when carrying out research. Names of bacteria and archaea are compiled in the List of Prokaryotic Names with Standing in Nomenclature (LPSN).
Taxonomists first tried to classify microbes as either animals or plants, which since ancient times had been considered the two “kingdoms” or major categories of life. For example, algae and fungi were included with plants. But German naturalist Ernst Haeckel (1834–1919) recognized that microbes differed from both plants and animals in fundamental aspects of their lifestyle, cell structure, and biochemistry. Haeckel proposed that microscopic organisms constitute a third kind of life—neither animal nor plant—which he called Monera.
In the twentieth century, biochemical studies revealed profound distinctions even within the Monera. In particular, microbes such as protists and algae contain a nucleus enclosed by a nuclear membrane, whereas bacteria do not. Herbert Copeland (1902–1968) proposed a system of classification that divided Monera into two groups: the Eukaryotes (protists, or protozoa and algae) and the Prokaryotes (bacteria). Copeland’s four-kingdom classification (plants, animals, protists, and prokaryotic bacteria) was later modified by Robert Whittaker (1920–1980) to include fungi as a fifth kingdom of eukaryotic microbes. But all these systems faced a challenge from the new science of genome sequences.
Archaea Differ from Bacteria and Eukaryotes
In 1977, Carl Woese (1928–2012), at the University of Illinois, was wondering how to measure the time since divergence of the most distantly related life forms. He recalled his earlier postdoctoral work on the coding of information in RNA molecules (Fig. 1.29B). Woese and a colleague, George Fox, used the sequence of the gene for 16S ribosomal RNA (16S rRNA) as a “molecular clock,” a gene whose sequence differences can be used to measure the time since the divergence of two species (discussed in Chapter 17). In constructing their tree of life, they included certain prokaryotes that live in seemingly hostile environments, such as the boiling sulfur springs of Yellowstone, or that conduct unusual kinds of metabolism, such as production of methane (methanogenesis). The divergence of rRNA genes showed that some of these prokaryotes were as distant from bacteria as they were from eukaryotes. These organisms represent a distinct domain of life: Archaea.
The archaea resemble the bacteria in their relatively simple cell structure and in their lack of a nucleus; thus, both archaea and bacteria are prokaryotes. But the gene expression machinery of archaea is more similar to that of eukaryotes. Archaea are found in a wide range of environments, and certain species, such as the autoclave-cultured archaeon Geogemma (Fig. 1.26B), grow in environments more extreme than any that support bacteria. Other kinds of archaea grow alongside bacteria in common soil or water—or even within the human gut or skin (discussed in Chapter 19).
Woese’s discovery replaced the classification scheme of five kingdoms with three equally distinct domains: Bacteria, Archaea, and Eukarya (Fig. 1.29A). But the genomes of undiscovered life had further surprises in store. In the twenty-first century, Jillian Banfield and co-workers at UC Berkeley pioneered the sequencing of metagenomes—the DNA sequences of entire microbial communities (Fig. 1.30). In 2004, Banfield’s research team sequenced the first environmental metagenome, which was obtained from a sample of extremely acidic mine drainage. Later environmental metagenomes revealed ultrasmall bacteria and archaea that can pass through a filter with 0.2-µm pores. Previously, the existence of such small cells was thought impossible.
More information
A photograph of Jillian Banfield sampling in a cave and a micrograph of an ultrasmall bacterium.
A photograph of Jillian Banfield sampling in a cave. Banfield wears long gloves, protective clothing, and a hard hat. A rusted pipe is visible above the collection site.
More information
A micrograph of an ultrasmall coccoid bacterium. The bacterium is 300 nanometers in diameter and lined with thin cilia.
FIGURE 1.30 ■Jillian Banfield sequences metagenomes.A. Banfield obtains samples from extremely acidic drainage at Richmond Mine, California. B. Ultrasmall bacterium obtained from a groundwater sample whose metagenomes revealed previously unknown clades of bacteria.
Source: Birgit Luef et al. 2015. Nat. Commun.6:6372.
MITCH JONES/JILLIAN BANFIELDB. LUEF ET AL. 2015. NAT. COMMUN. 6:6372
Eukaryotes Evolved through Endosymbiosis
During the early twentieth century, microscopists wondered about the nature of subcellular structures such as mitochondria and chloroplasts, which have the appearance of cells within cells. Lynn Margulis (1938–2011) at the University of Massachusetts was fascinated by organelles amid the diversity of microbial eukaryotes (Fig. 1.31). Margulis tried to explain why eukaryotic cells contain mitochondria and chloroplasts, membranous organelles that possess their own chromosomes. She proposed that eukaryotes evolved by merging with bacteria to form composite cells by intracellular endosymbiosis, in which one cell internalizes another that grows within it. The endosymbiosis may ultimately generate a single organism whose formerly independent members are now incapable of independent existence.
More information
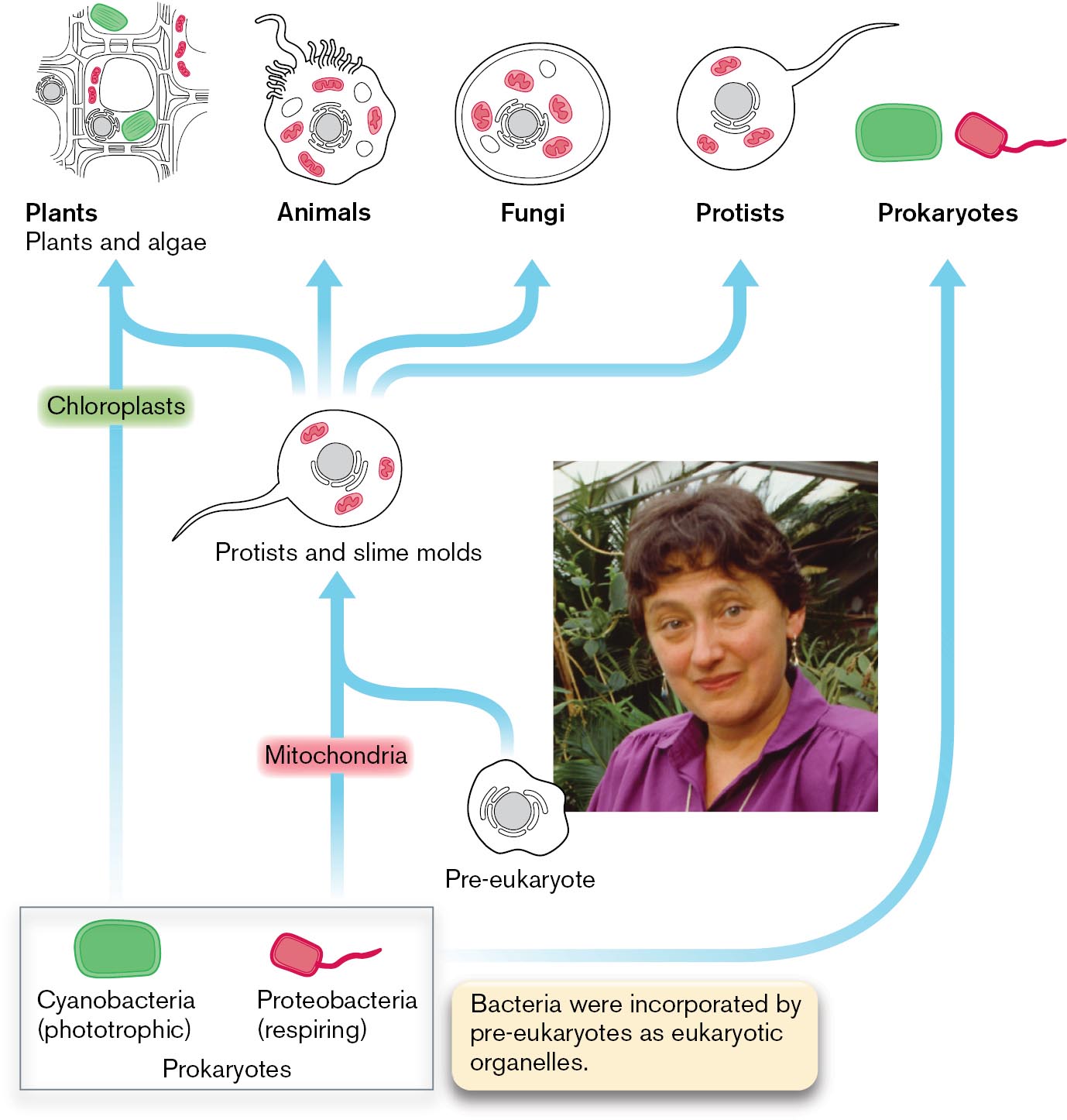
An illustration of the theory of serial endosymbiosis with an inset photo of Lynn Margulis. The illustration shows the structures of cells such as plants and algae, animals, fungi, protists, and prokaryotes in a row. The prokaryotes symbol is further broken down into two images of Cyanobacteria, labeled phototrophic, and Proteobacteria, labeled respiring. The corresponding text reads bacteria were incorporated by pre-eukaryotes as eukaryotic organelles. An arrow links Cyanobacteria to plants and algae through chloroplasts. Chloroplasts are highlighted within the drawing of the plant cell structure. An arrow links Proteobacteria to protists and slime molds through mitochondria and a pre-eukaryote. Mitochondria are highlighted within the drawing of a protist. Arrows then link the protist to plants, animals, fungi, and protists. Mitochondria are highlighted in the cell structures of plants, animals, fungi, and protists.
An electron micrograph of Chlorobium species. The bacterium has a circular shape, a few structures are labeled. The outer layer is labeled as the envelope. A labeled nucleoid is partially visible in the center. Oval-shaped structures that sit close to the cell membrane are labeled as chlorosomes. The bacterium is about 400 nanometers in diameter.
FIGURE 1.31 ■Lynn Margulis and the serial endosymbiosis theory.Inset: Lynn Margulis proposed that organelles evolve through endosymbiosis.NANCY R. SCHIFF/GETTY IMAGES
Margulis proposed that early in the history of life, respiring bacteria similar to Escherichia coli were engulfed by pre-eukaryotic cells, where they evolved into mitochondria, the eukaryote’s respiratory organelles. Similarly, she proposed that a phototroph related to cyanobacteria was taken up by a eukaryote, giving rise to the chloroplasts of phototrophic algae and plants. In Margulis’s model, the nature of the original pre-eukaryote that took up precursors to mitochondria was unclear. Today, genomic evidence suggests that the original pre-eukaryote may have been an archaeon (discussed in Chapter 19).
The endosymbiosis theory was controversial because it implied a polyphyletic, or multiple, ancestry of living species, inconsistent with the long-held assumption that species evolve only by divergence from a common ancestor (monophyletic ancestry). Ultimately, DNA sequence analysis produced compelling evidence of the bacterial origin of mitochondria and chloroplasts. Both of these classes of organelles contain circular molecules of DNA, whose sequences show unmistakable homology (similarity) to those of bacteria. Genome sequences now reveal many cases of horizontal gene transfer among all three domains of life.
Thought Question
1.10 What arguments support the classification of Archaea as a third domain of life? What arguments support the classification of archaea and bacteria together, as prokaryotes, distinct from eukaryotes?
ANSWER ANSWER
The sequence of 16S rRNA (small-subunit rRNA) and other fundamental genes differs as much between archaea and bacteria as it does between archaea and eukaryotes. The composition of archaeal cell walls and phospholipids is completely distinct from that of bacteria and eukaryotes. Some aspects of gene expression, such as the RNA polymerase complex, are more similar between archaea and eukaryotes than between archaea and bacteria. On the other hand, archaeal and bacterial cells are prokaryotic; they both lack nuclei and complex membranous organelles. Archaeal metabolism and lifestyles are more similar to those of bacteria than to those of eukaryotes. Some archaea and bacteria sharing the same environment, such as high-temperature springs, have undergone horizontal transfer of genes that encode traits such as heat-stable membrane lipids.
To Summarize
Classifying microbes was a challenge historically because of the difficulties in observing distinguishing characteristics of different categories.
Ernst Haeckel recognized that microbes constitute a form of life distinct from animals and plants.
Herbert Copeland and Robert Whittaker classified prokaryotes as a form of microbial life distinct from eukaryotic microbes such as protists.
Carl Woese discovered a domain of prokaryotes, Archaea, whose genome sequences diverge equally from those of bacteria and those of eukaryotes. Archaea grow in a wide range of environments; some species grow under conditions that exclude bacteria and eukaryotes.
Jillian Banfield pioneered the sequencing of metagenomes that reveal previously unknown forms of microbial life.
Lynn Margulis proposed that eukaryotic organelles such as mitochondria and chloroplasts evolved by endosymbiosis from prokaryotic cells engulfed by pre-eukaryotes.
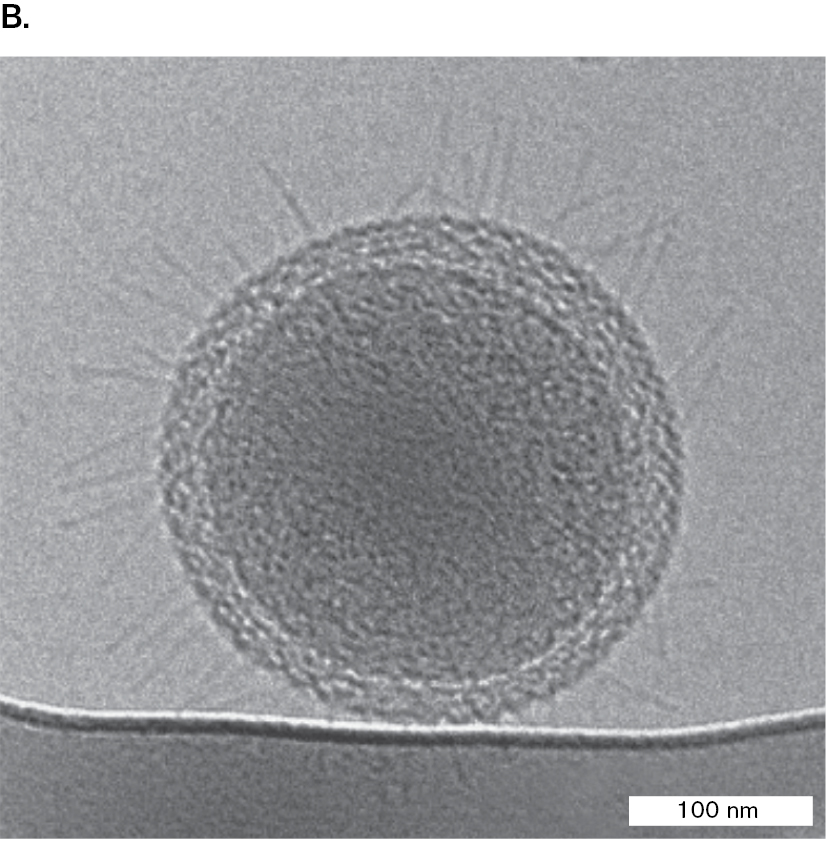
Image B is an electron micrograph showing Geogemma strain 121, and iron oxide. Geogemma is a large round archaeon with many thin flagella protruding from a single point. Geogemma has a diameter of 1.5 micrometers. Next to Geogemma, there is a buildup of iron oxide.
B.Geogemma barossii is a round archaeon with a tuft of flagella (transmission electron micrograph). K. KASHEFI AND D. R. LOVLEY. 2003. SCIENCE 301:934


 ANSWER
ANSWER ANSWER
ANSWER