The form and function of a membrane depends upon its lipids and proteins (see Fig. 3.2). The most common type of membrane lipids are phospholipids (Fig. 3.5). Bacterial membranes consist of a bilayer of lipids that face each other tail to tail, keeping their hydrophobic side chains away from the water inside and outside the cell. The two layers of phospholipids in the bilayer are called leaflets. One leaflet of phospholipids faces the cell interior; the other faces the exterior. As a whole, the phospholipid bilayer imparts fluidity and gives the membrane a consistent thickness (about 8 nm).
More information
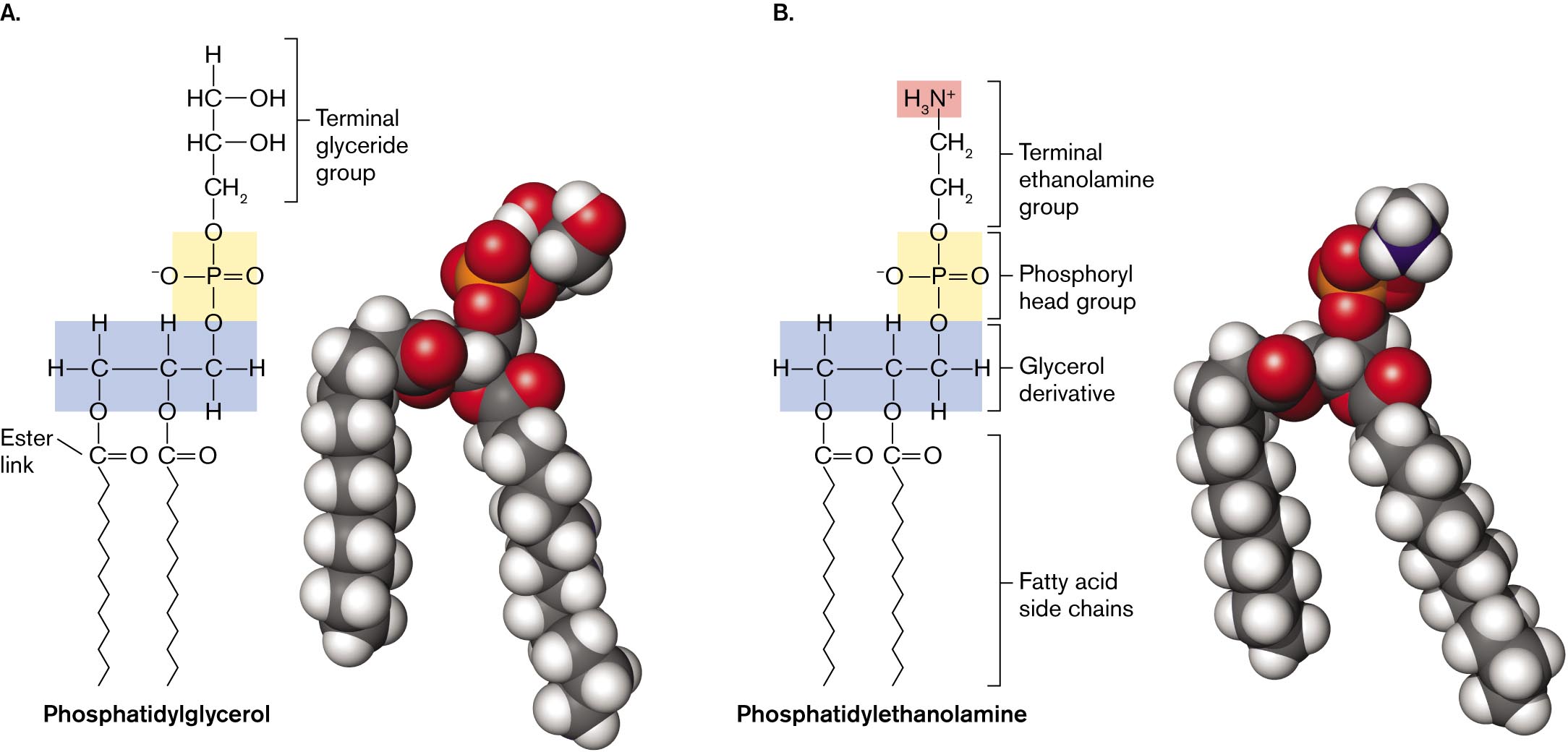
Two molecular structures of phospholipids with accompanying space filling models.
The molecular structure and space filling model of phosphatidylglycerol. The molecular structure consists of a,3 membered carbon chain, in which the left carbon is single bonded to hydrogen at the top and left, single bonded to oxygen at the bottom. Carbon in the center is single bonded to hydrogen at the top and single bonded to oxygen at the bottom. The carbon at the right is single bonded to a phosphate group at the top and single bonded to hydrogen at the bottom and right. The phosphate group consists of phosphorous in the center single bonded to oxygen at the top and bottom, single bonded to oxygen minus ion at the left, and double-bonded to oxygen at the right. The terminal glyceride group is attached to the oxygen of the phosphate, which consists of H C in the center single bonded to hydrogen at the top and single bonded to H C at the bottom. Both the H C groups are single bonded to OH groups at the right. H C at the bottom is single bonded to methyl. The oxygens bonded to the left and center carbon are each single bonded to carbon double-bonded to oxygen at the right labeled as ester link, which is attached to a carbon chain at the bottom. A space-filling model of Phosphatidylglycerol is shown at the right. In the model, carbon atoms are black spheres, hydrogen atoms are white spheres, and oxygen atoms are red spheres.
The molecular structure and space filling model of phosphatidylethanolamine. The molecular structure consists of a 3 membered carbon chain, in which the left carbon is single bonded to hydrogen at the top and left, single bonded to oxygen at the bottom. Carbon in the center is single bonded to hydrogen at the top and single bonded to oxygen at the bottom. The carbon at the right is single bonded to the phosphoryl head group at the top, single bonded to hydrogen at the right, and single bonded to hydrogen at the bottom. The phosphate group consists of phosphorous in the center is single bonded to oxygen at the top and bottom, single bonded to oxygen minus ion at the left, and double-bonded to oxygen at the right. The terminal ethanolamine group is attached to the oxygen of the phosphate, and consists of a C H subscript 2 bound to a C H subscript 2 bound to H subscript 2 N plus ion at the top. The oxygens bonded to the left and center carbon are single bonded to carbon double-bonded to oxygen at the right, in which the carbons are attached to the fatty acid side chain at the bottom. A space-filling model of Phosphatidylethanolamine is shown. In the model, carbon atoms are black spheres, hydrogen atoms are white spheres, and oxygen atoms are red spheres.
FIGURE 3.5 ■Phospholipids.A. Phosphatidyl glycerolconsists of glycerol with ester links to two fatty acids, and a phosphoryl group linked to a terminal glyceride. B. Phosphatidyl ethanolaminecontains a glycerol linked to two fatty acids, and a phosphoryl group with a terminal ethanolamine. The ethanolamine carries a positive charge.
Membrane Lipids
A phospholipid possesses a charged phosphoryl “head” that contacts the water interface, as well as a hydrophobic “tail” of fatty acids packed within the bilayer. Lipid biosynthesis is a key process that is vulnerable to some antibiotics. For example, the bacterial enzyme enoyl reductase, which synthesizes fatty acids (discussed in Chapter 15), is the target of triclosan, a common antibacterial additive in detergents and cosmetics.
A typical phospholipid consists of glycerol with ester links to each of two fatty acids, and a phosphoryl polar head group, which at neutral pH is deprotonated (negatively charged; Fig. 3.5). This kind of phospholipid is called a phosphatidate. The negatively charged head group of the phosphatidate can contain various organic groups, such as glycerol to form phosphatidylglycerol (Fig. 3.5A). In other lipids, the polar head group has a side chain with positive charge. The positive charge commonly resides on an amine group, such as ethanolamine in phosphatidylethanolamine (Fig. 3.5B). Phospholipids with positive charge or with mixed charges are concentrated in portions of the membrane that interact with DNA, which has negative charge.
Membranes require a uniform thickness and stability to maintain structural integrity and function. So why do individual membrane lipids differ in structure? Different environments favor different forms of membrane lipids. For example, lipid structure helps determine whether an organism can grow in a hot spring or whether it can colonize human lungs.
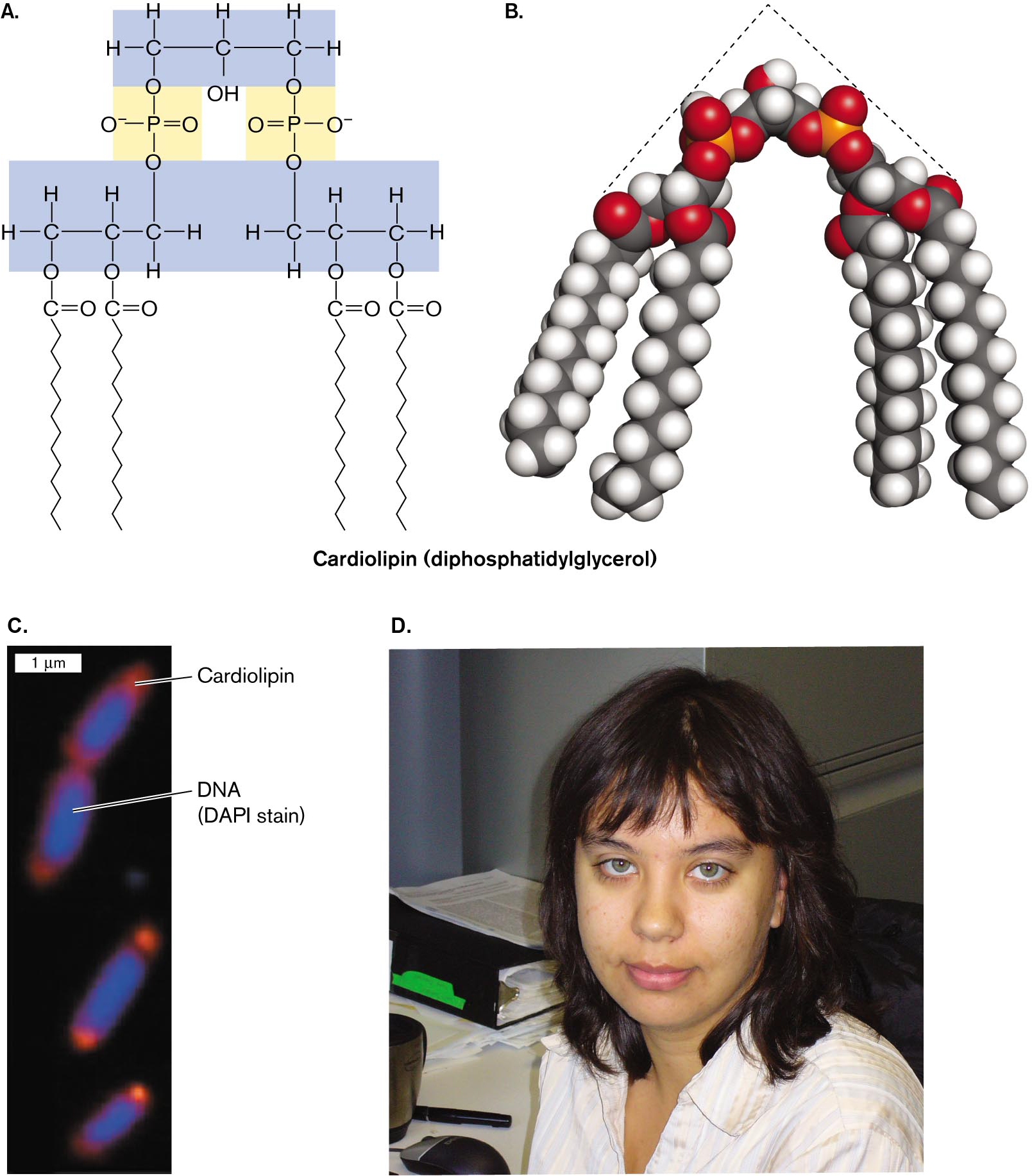
Environmental stress. Starvation stress increases bacterial production of lipids with an unusual type of phosphoryl head group. Cardiolipin, or diphosphatidylglycerol, is actually a double phospholipid linked by a glycerol (Fig. 3.6). Cardiolipin concentration increases in bacteria grown to starvation or stationary phase (discussed in Chapter 4). Tatyana Romantsov and Janet Wood, at the University of Guelph, showed how cardiolipin helps define the polar structure of a bacterial cell.
Within a rod-shaped cell, cardiolipin does not diffuse at random; it concentrates in patches called “domains” near the cell poles. The polar localization of cardiolipin was demonstrated by fluorescence microscopy, in which a cardiolipin-specific fluorophore localized to the poles of Escherichia coli. The “wedge” shape of cardiolipin (Fig. 3.6B), with its narrow head group and wide fatty acid group, is thought to form concave domains of lipid that stabilize the curve of the polar membrane. Cardiolipin may enhance the formation of smaller cells during starvation. At the cell pole (Fig. 3.6C), cardiolipin binds certain environmental stress proteins, such as a protein that transports osmoprotectants when the cell is under osmotic stress. Thus, a phospholipid can have specific functions associated with specific membrane proteins.
More information
The molecular structure, a space filling model, and a micrograph of cardiolipin, accompanied by a photo of Tatyana Romantsov.
The molecular structure of cardiolipin. The cardiolipin structure consists of a glycerol group with two phospholipids attached. The glycerol group consists of a three carbon chain, in which C 1 and C 3 are each single bonded to the oxygens of phosphate groups, single bonded to two hydrogens, and single bonded to C 2. C 2 is single bonded to a hydrogen, single bonded to a hydroxyl group, single bonded to C 1 and C 3. The phosphate groups make up the heads of the phospholipids. Each phosphate group consists of a central phosphorus atom single bonded to two oxygens, single bonded to a negatively charged oxygen atom, and double bonded to an oxygen. In each phosphate group, one of the single bonded oxygen atoms connects to a carbon of the glycerol and the other single bonded oxygen atom connects to a carbon of the phospholipid tail structure. The phospholipid tail structures each consist of a three carbon chain, in which C 1, or C 3, is single bonded to oxygen of the phosphate group. This carbon is also single bonded to two hydrogen atoms and single bonded to C 2 of the carbon chain. C2 of the carbon chain is single bonded to a hydrogen and single bonded to an oxygen. This oxygen is single bonded to a carbon that is double bonded to an oxygen and single bonded to a long carbon chain. C 3 or C 1 of the three carbon chain is single bonded to two hydrogen atoms and single bonded to an oxygen. This oxygen is single bonded to a carbon atom that is double bonded to an oxygen and single bonded to a long carbon chain.
A space-filling model of cardiolipin. The model shows the three-carbon group as a point to which two phospholipids are attached. The phospholipids are angled from the glycerol group in a triangular shape. In the model, carbon atoms are represented by black spheres, hydrogen atoms are represented by white spheres, oxygen atoms are represented by red spheres, and phosphorus atoms are represented by orange spheres.
An electron micrograph of cardiolipin pole localization in bacterial cells. Four rod shaped bacterial cells are shown, each about 1 micrometer long and 0.5 micrometer wide. The cardiolipin is visible at both ends of each cell. In the center of the cell D N A is visible. The cardiolipin appears orange due to a specialized fluorophore, and the D N A appears blue due to D A P I stain.
A photo of Tatyana Romantsov. Tatyana has brown hair and blue eyes. She is smiling at the camera in the photo.
FIGURE 3.6 ■Cardiolipin localizes to the poles.A. Cardiolipin is a double phospholipid joined by a third glycerol. B. A space-filling model of cardiolipin shows its triangular shape. C. Cardiolipin localizes to the bacterial cell poles, as shown by microscopy with a cardiolipin-specific fluorophore (orange). D. Tatyana Romantsov, at the University of Guelph, investigates how cardiolipin helps form the shape of a cell pole. TATYANA ROMANTSOV ET AL. 2007. MOL. MICROBIOL.64:1455TATYANA ROMANTSOV
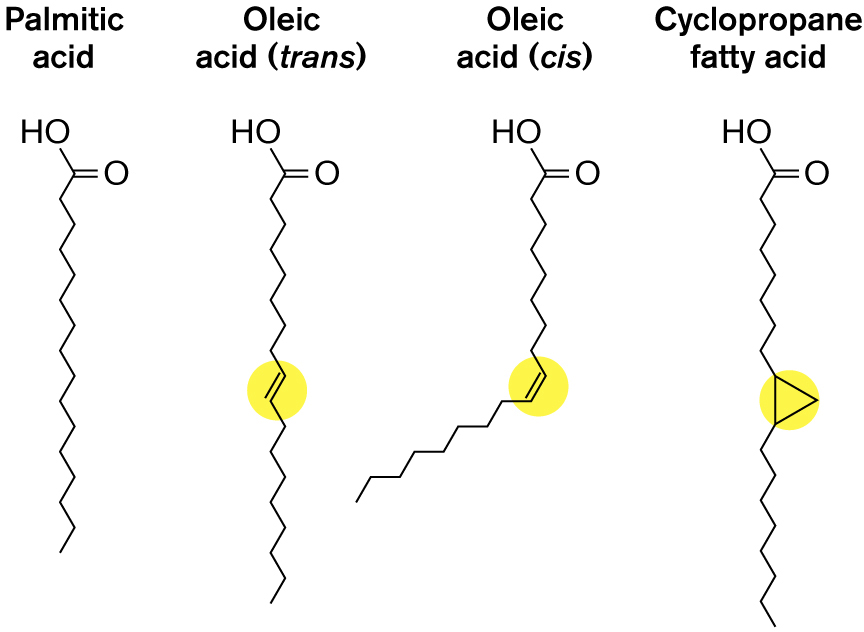
The fatty acid component of phospholipids also varies. The most common bacterial fatty acids are hydrogenated chains of varying length, typically between 6 and 22 carbons. But some fatty acid chains are partly unsaturated (possess one or more carbon-carbon double bonds). Most unsaturated bonds in membranes are cis, meaning that both alkyl chains are on the same side of the bond, so the unsaturated chain has a “kink,” as in the cis form of oleic acid (Fig. 3.7). Because the kinked chains do not pack as closely as the straight hydrocarbon chains do, the membrane is more “fluid.” This is why, at room temperature, unsaturated vegetable oils are fluid, whereas highly saturated butterfat is solid. The enhanced fluidity of a kinked phospholipid improves the function of the membrane at low temperature; hence, bacteria can respond to cold and heat by increasing or decreasing their synthesis of unsaturated phospholipids.
More information
The molecular structures of four phospholipid side chains are shown. The four side chains are palmitic acid, oleic acid in a trans formation, oleic acid in a cis formation, and cyclopropane fatty acid. The palmitic acid consists of a 16-membered carbon chain, in which C 1 is attached with hydroxyl, and C 2 is double-bonded to oxygen at the right. The oleic acid, trans formation, shows an 18 membered carbon chain in which C 1 is attached with hydroxyl, and C 2 is double-bonded to an oxygen at the right. C 10 and C 11 in the chain are double-bonded. The oleic acid, cis formation, shows an 18 membered carbon side chain, in which C 1 is attached to hydroxyl, and C 2 is double bonded to oxygen. C 10 and C 11 in the chain are double bonded and the direction of the chain is diverted by this bond. The Cyclopropane fatty acid consists of a 20-membered carbon chain in which C 1 is attached with hydroxyl, and C 2 is double-bonded to oxygen at the right, and C 10 and C 11 in the chain form a triangle.
FIGURE 3.7 ■Phospholipid side chains.
Another interesting structural variation is cyclization of part of the chain to form a stiff planar ring with decreased fluidity. The double bond of unsaturated fatty acids can incorporate a carbon from S-adenosyl-l-methionine to form a three-membered ring, generating a cyclopropane fatty acid (Fig. 3.7). Bacteria convert unsaturated fatty acids to cyclopropane during starvation and acid stress, conditions under which membranes require stiffening. Cyclopropane conversion is an important factor in the pathogenesis of Mycobacterium tuberculosis and in the acid resistance of food-borne toxigenic E. coli.
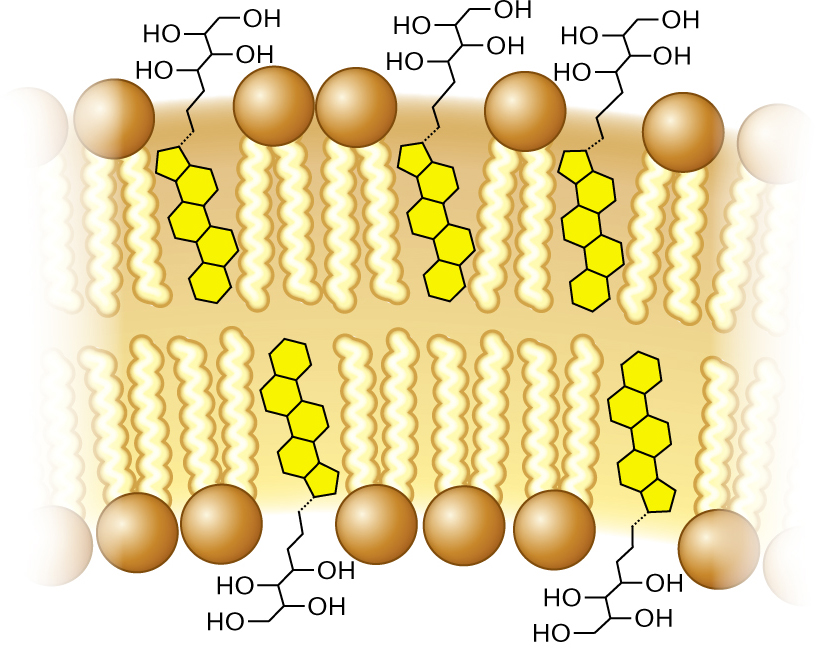
In addition to phospholipids, membranes include planar molecules that fill gaps between hydrocarbon chains. These stiff, planar molecules reinforce the membrane, much as steel rods reinforce concrete. In eukaryotic membranes, the reinforcing agents are sterols, such as cholesterol. In some bacteria, the same function is filled by pentacyclic (five-ring) hydrocarbon derivatives called hopanoids, or hopanes (Fig. 3.8). Like cholesterol, hopanoids fit between the fatty acid side chains of membranes and limit their motion, thus stiffening the membrane. Hopanoids provide biomarkers for petroleum exploration, as signs of potential petroleum formation in ancient rock.
More information
A detailed illustration of cell membrane phospholipids and hopanoids. The phospholipid bilayer of the cell membrane is visible. Phospholipid tails are located interiorly. Hopanoids are present between every few phospholipids. The hopanoid structure consists of five benzene rings fused together and single bonded to an eight carbon chain. The first four carbons in the chain are each single-bonded to a hydroxyl group. The carbon chain of the hopanoid is located to the exterior of the membrane and the benzene ring structures are located in the interior of the membrane.
FIGURE 3.8 ■Hopanoids add strength to membranes. Hopanoids limit the motion of phospholipid tails, thus stiffening the membrane.
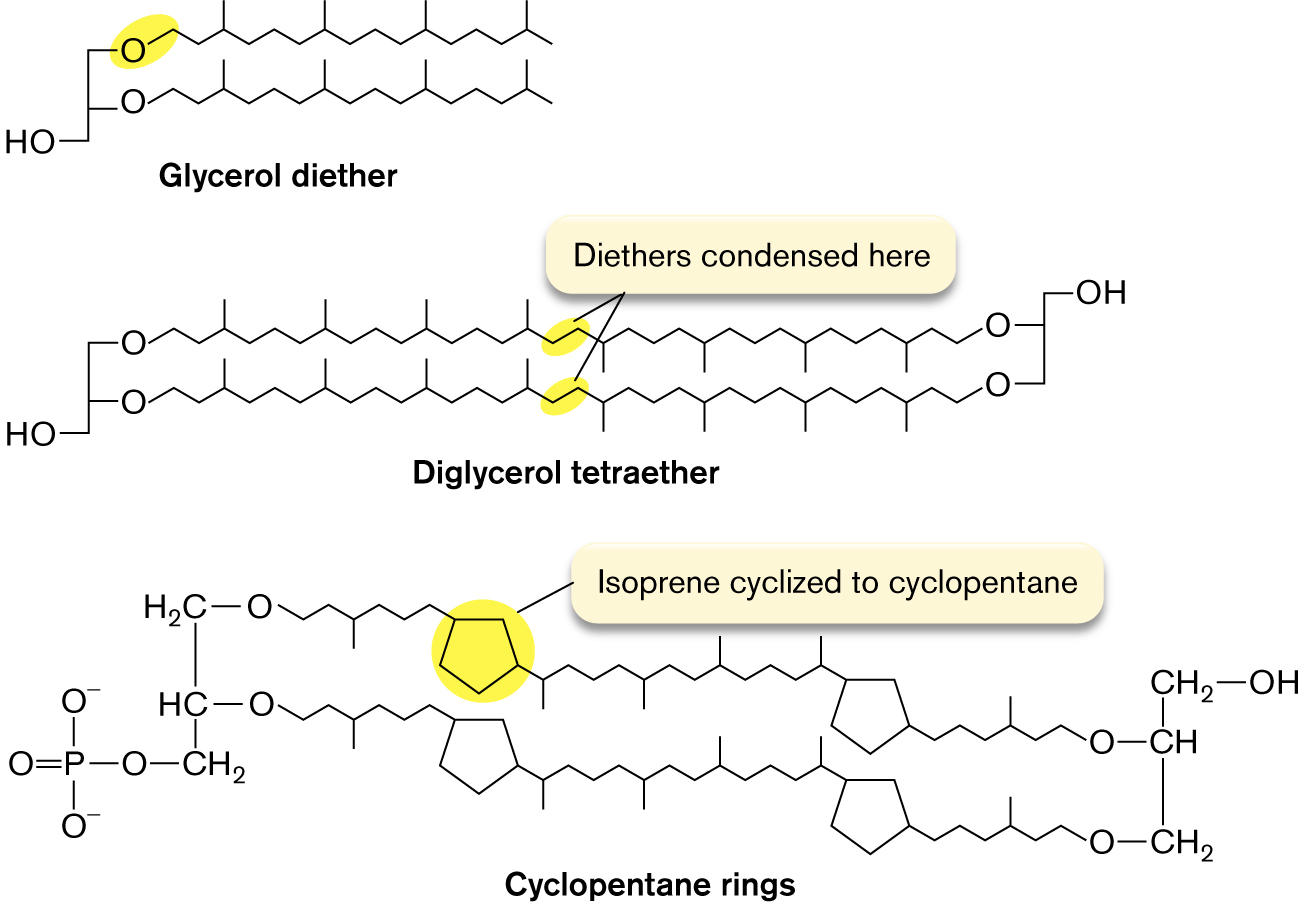
Archaea have unique membrane lipids. The membrane lipids of archaea differ fundamentally from those of bacteria and of eukaryotes. All archaeal phospholipids replace the ester link between glycerol and fatty acid with an ether link, C–O–C (Fig. 3.9). Ethers are much more stable than esters, which hydrolyze easily in water. This is one reason why some archaea can grow at higher temperatures than all other forms of life. Another modification is that archaeal hydrocarbon chains are branched terpenoids, polymeric structures derived from isoprene, in which every fourth carbon extends a methyl branch. The branches strengthen the membrane by limiting movement of the hydrocarbon chains.
More information
The molecular structures of glycerol diether, diglycerol tetraether, and cyclopentane rings are shown. Glycerol diether consists of an O H group single bonded to two oxygens. Each oxygen is bonded to a 16 member carbon chain. Diglycerol tetraether consists of an O H group single bonded to two oxygens. Each oxygen is bonded to a 20 member carbon chain. The final carbon in each chain is bonded to an oxygen. These two oxygens are bonded to an O H group. A note at the middle of the carbon chains reads, diethers condensed here.
The structure of Cyclopentane rings consists of 2 parallel long carbon chains bonded with two 5-membered carbon rings in the left and the right. C 1 of the first chain is attached with oxygen is single bonded to H 2 C. C 1 of the second carbon chain is attached with oxygen is single bonded to H C. The carbon in the first chain is single bonded to carbon in the second chain, further carbon is single bonded to methyl, and the methyl is single bonded to a phosphate group. The last carbon chain of the C 1 chain is single bonded to oxygen, and the oxygen is single bonded to C H, C H is single bonded to methyl in which methyl is single bonded to the hydroxyl group at the right. The last chain of the C 2 carbon chain is attached to oxygen, and the oxygen is single bonded to methyl, in which methyl is single bonded to C H in the first chain. The five-membered ring on the left of the first carbon chain is shown highlighted, and the text reads Isoprene cyclized to cyclopentane.
FIGURE 3.9 ■Terpene-derived lipids of archaea. In archaea, the hydrocarbon chains are ether-linked to glycerol, and every fourth carbon has a methyl branch. In some archaea, the tails of the two facing lipids of the bilayer are fused, forming tetraethers; thus, the entire membrane consists of a monolayer.
The most extreme hyperthermophiles, which live beneath the ocean at 110°C, have terpenoid chains linked at the tails, forming a tetraether monolayer. In some species, the terpenoids cyclize to form cyclopentane rings. These planar rings stiffen the membrane under stress to an even greater extent than do the cyclopropane chains of bacteria. (Thermophiles and other microbes in extreme environments are presented in Chapter 5. For more on archaea, see Chapter 19.)
Membrane Proteins
Membrane proteins are diverse and can serve different functions. Different membrane proteins provide structural support, communicate with the environment, secrete virulence factors, and mediate transport. Certain proteins have very specific roles; for example, the TetA protein expels the antibiotic tetracycline, making a pathogen resistant to tetracycline.
Structural support. Some membrane proteins anchor together different layers of the cell envelope (discussed in Section 3.3). Other proteins attach the membrane to the cytoskeleton or form the base of structures extending out from the cell, such as flagella.
Detection of environmental signals. In Vibrio cholerae, the causative agent of cholera, the membrane protein ToxR detects acidity and elevated temperature—signs that the bacterium is in the host’s digestive tract. The ToxR domain facing the cytoplasm then binds to a DNA sequence, activating expression of cholera toxin.
Secretion of virulence factors and communication signals. Membrane protein complexes export toxins and cell signals across the envelope. For example, symbiotic nitrogen-fixing rhizobia require membrane proteins NodI and NodJ to transport nodulation signals out to the host plant roots, inducing the plant to form root nodules containing the bacteria.
Ion transport and energy storage. Transport of ions across a membrane generates a transmembrane gradient that stores energy.
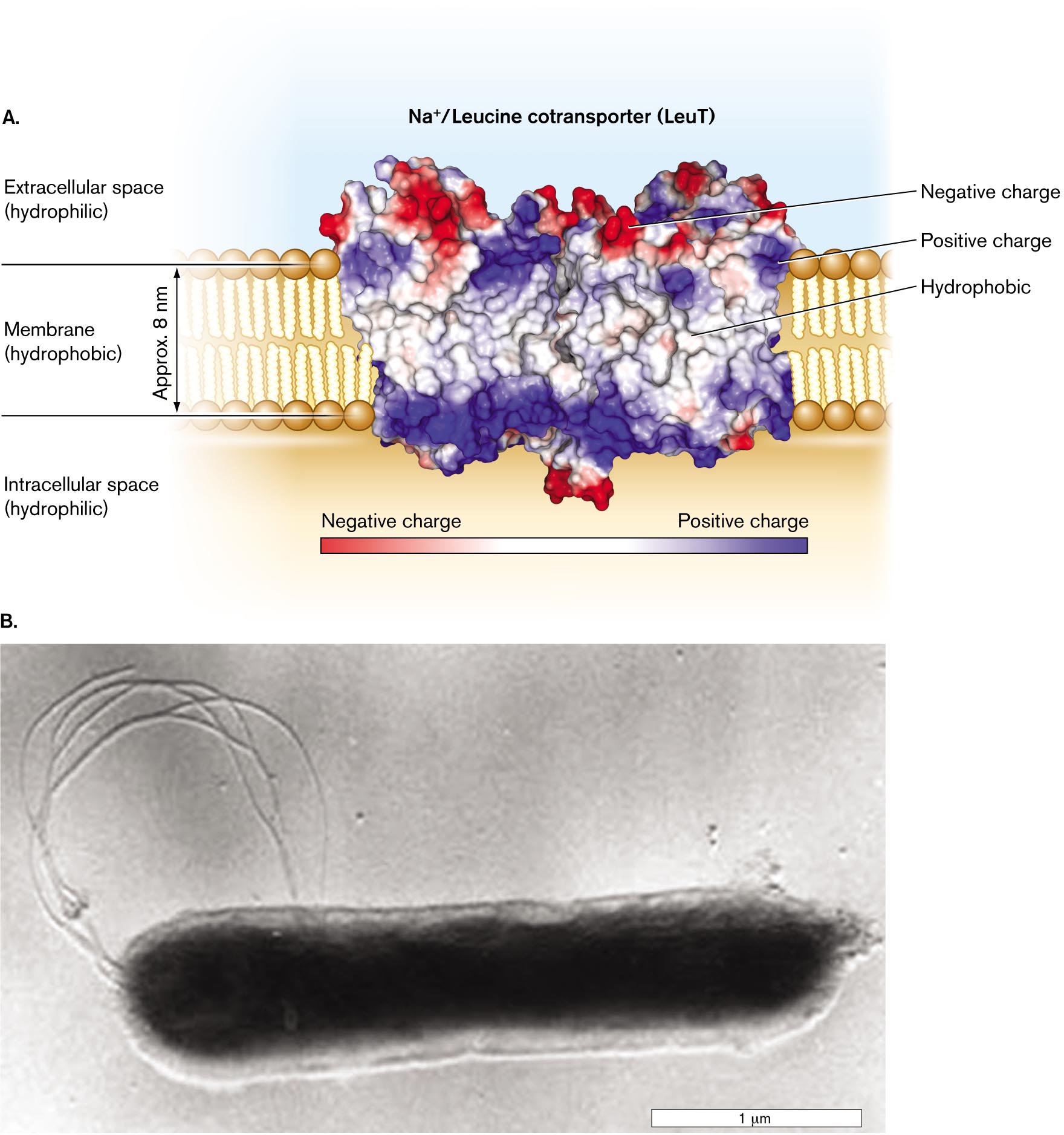
An example of a membrane protein is the leucine transporter LeuT (Fig. 3.10A). LeuT drives uptake of leucine, coupled to a gradient of sodium ions. The protein complex was purified for X-ray diffraction from Aquifex aeolicus, a thermophilic bacterium whose heat-stable proteins form durable crystals (Fig. 3.10B). Remarkably, LeuT is homologous (shares common ancestry) with a human neuron protein that transports neurotransmitters. Thus, this bacterial protein serves as a model for the study of neuron function.
LeuT and other proteins embedded in a membrane require a portion of hydrophobic amino-acid side chains that are soluble amidst the hydrocarbon tails of the phospholipids. (For a review of protein structure, see eAppendix 1.) The hydrophobic portions of LeuT are shown in white in Figure 3.10A. Typically, several hydrophobic alpha helices thread back and forth through the membrane. Their middle surfaces, deeply embedded in membrane, have hydrophobic amino acid residues that interact favorably with the hydrocarbon chains of the phospholipids. Other peptide regions extend outside the membrane, containing charged and polar amino acids that interact favorably with the negatively charged head groups (such as phosphoryl groups) and with water.
Figure 3.10A also shows the LeuT charge distribution. Hydrophobic amino acid residues (white) make the protein soluble in the membrane, while portions with negative charge (red) and positive charge (blue) interact with the phospholipid head groups and with water. The net result of these interactions is to lock the protein securely within the membrane.
More information
A detailed illustration of the LeuT sodium/leucine cotransporter and a micrograph of the bacterium Aquifex aeolicus.
A detailed illustration of the LeuT sodium/leucine cotransporter in the cell membrane. The protein is an irregular rectangular shape and wider than the cell membrane. The cell membrane is identified as a hydrophobic region of about 0.8 nanometer wide. The space exterior to the membrane outside of the cell is labeled as the extracellular space and identified as a hydrophilic region. The space exterior to the membrane within the cell is labeled as the intracellular space and identified as a hydrophilic region. The protein has regions that are negatively charged and regions that are positively charged. Most negatively charged regions are oriented towards the extracellular space. Most positively charged regions are oriented towards the intracellular space. The region of the protein located within the membrane is labeled hydrophobic and has a neutral charge.
A micrograph of the bacterium Aquifex aeolicus. The bacterium is rod shaped and has flagella. The cell is about 2.5 micrometers long and 0.5 micrometer wide. Internal structures are not visible in the micrograph.
FIGURE 3.10 ■A cell membrane–embedded transport protein: the LeuT sodium/leucine cotransporter of Aquifex bacteria.A. The protein complex carries leucine across the cell membrane into the cytoplasm, coupled to sodium ion influx. (PDB code: 3F3E) B.Aquifex aeolicus grows at 96°C in hot springs. KARL STETTER AND REINHARDT RACHEL, U. REGENSBURG, GERMANY
Molecules Cross the Cell Membrane
The cell membrane acts as a barrier to keep water-soluble proteins and other cell components within the cytoplasm. But how do nutrients from outside get into the cell—and how do secreted products such as toxins get out? Specific membrane proteins transport molecules across the membrane between the cytoplasm and the outside. Selective transport is essential for cell survival; it means the ability to acquire scarce nutrients, exclude waste, and transmit signals to neighbor cells.
Passive diffusion. Small, uncharged molecules, such as O2, CO2, and water, easily permeate the membrane. Some molecules, such as ethanol, also disrupt the membrane—an action that can make such molecules toxic to cells. By contrast, large, strongly polar molecules such as sugars, and charged molecules such as amino acids, generally cannot penetrate the hydrophobic interior of the membrane and thus require transport by specific proteins. Water molecules permeate the membrane, but their rate of passage is increased by protein channels called aquaporins (discussed in Chapter 4).
Osmosis. Most cells maintain a concentration of total solutes (molecules in solution) that is higher inside the cell than outside. As a result, the internal concentration of water is lower than the concentration outside the cell. Because water can cross the membrane but charged solutes cannot, water tends to diffuse across the membrane into the cell, causing the expansion of cell volume, in a process called osmosis. The resulting pressure on the cell membrane is called osmotic pressure, or turgor pressure (see Figure A2.5 in eAppendix 2). Osmotic pressure will cause a cell to burst, or lyse, in the absence of a countering pressure such as that provided by the cell wall. That is how penicillin kills bacteria—by disrupting cell wall synthesis.
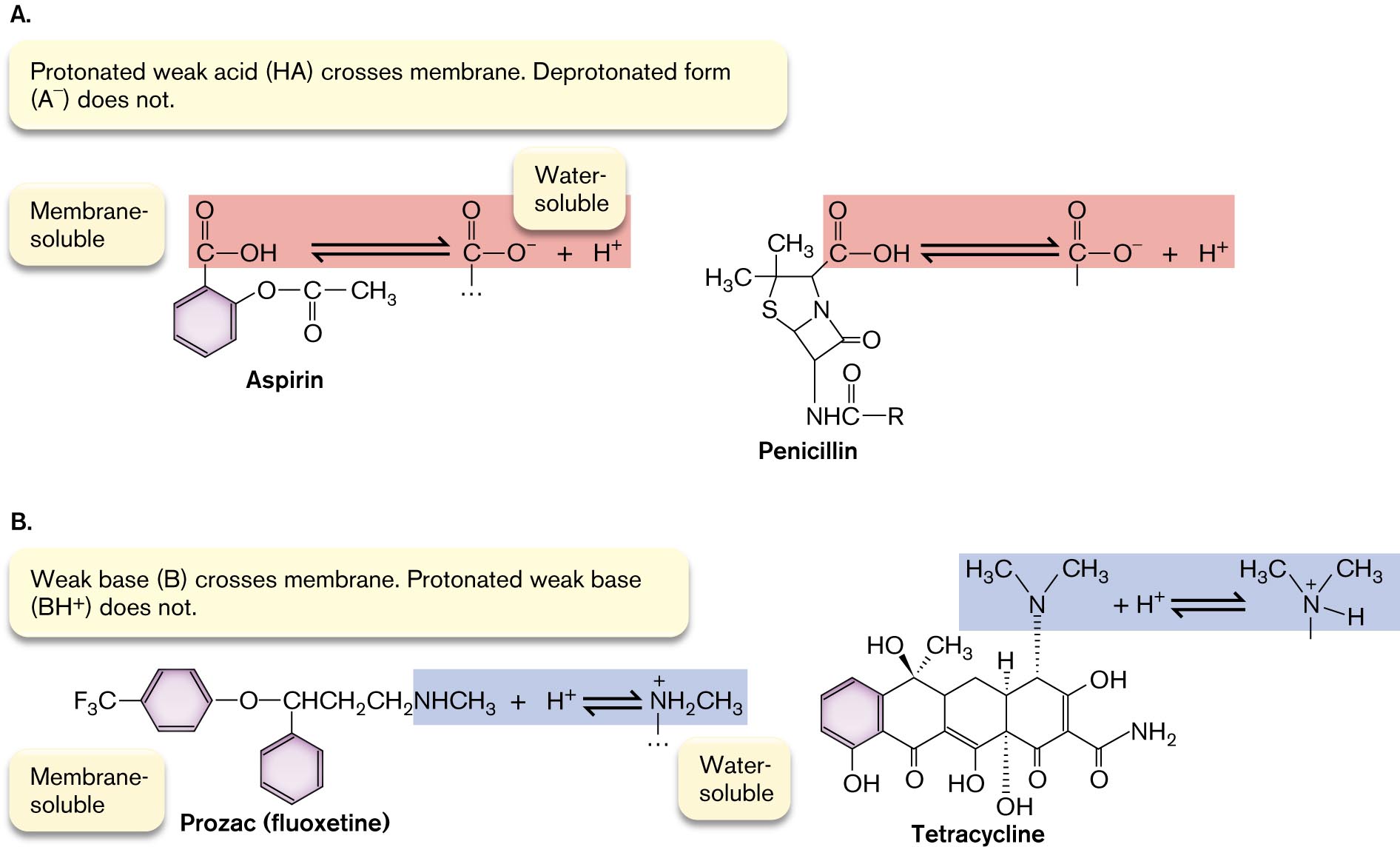
Membrane-permeant weak acids and weak bases. A special case of movement across cell membranes is that of membrane-permeant weak acids and membrane-permeant weak bases (Fig. 3.11), which exist in equilibrium between charged and uncharged forms:
Weak acid: HA ⇌ H+ + A−
Weak base: B + H2O ⇌ BH+ + OH−
More information
The molecular structures of Aspirin, Penicillin, Prozac, and Tetracycline are shown.
Membrane soluble and water soluble molecular structures of Aspirin and Penicillin. The text reads, Protonated weak acid, H A, crosses membrane. Deprotonated form, A superscript minus, does not. The molecular structures of membrane soluble and water soluble Aspirin are shown. The core structure of Aspirin consists of a benzene ring single bonded to an oxygen atom. The oxygen is single bonded to a carbon that is double bonded to an oxygen and single bonded to a C H subscript 3 group. In Aspirin’s membrane soluble form, the benzene ring is single bonded to a carbon that is double bonded to an oxygen and single bonded to an O H group. In Aspirin’s water soluble form, the benzene ring is single bonded to a carbon that is double bonded to an oxygen atom and single bonded to a negatively charged oxygen ion. This is shown in reaction with H superscript plus. The molecular structures of membrane soluble and water soluble Penicillin are shown. The core structure of Penicillin consists of a four membered ring fused to a five membered ring. In the four membered ring, C 1 is double bonded to oxygen, C 2 is single bonded to a nitrogen, and C 4 is replaced with a nitrogen. The C 2 nitrogen is single bonded to hydrogen and carbon. The carbon is double bonded to oxygen and single bonded to an R group. In the five membered ring, C 2 is replaced with a nitrogen, C 4 is replaced with a sulfur atom, and C 5 is single bonded to two C H subscript 3 groups. In Penicillin’s membrane soluble form, C 1 of the five membered ring is single bonded to a carbon that is double bonded to oxygen and single bonded to an O H group. In Penicillin’s water soluble form, C 1 of the five membered ring is single bonded to a carbon that is double bonded to oxygen and single bonded to a negatively charged oxygen ion in reaction with H superscript plus.
Membrane soluble and water soluble molecular structures of Prozac, or fluoxetine, and Tetracycline. The text reads, weak base, B, crosses membrane. Protonated weak base, B H superscript plus, does not. The membrane soluble and water soluble structures of Prozac are shown. The core structure consists of two benzene rings. In the first ring, C 1 is single bonded to oxygen that is single bonded to C H C H subscript 2 C H subscript 2. In the second ring, C 1 is single bonded to the same C H C H subscript 2 C H subscript 2 group as the first ring. In Prozac’s membrane soluble form, the C H C H subscript 2 C H subscript 2 group is bounded to N H C H subscript 3 in reaction with H superscript plus. In Prozac’s water soluble form, the C H C H subscript 2 C H subscript 2 group is bounded to N plus H subscript 2 C H subscript 3. The membrane soluble and water soluble structures of Tetracycline are shown. The core structure of Tetracycline consists of four benzene rings fused together. In the first ring, C 1 is single bonded to O H. In the second ring, C 1 is double bonded to O and C 4 is single bonded to O H at angle closer to the viewer and single bonded to C H subscript 3 at angle farther from the viewer. In the third ring, C 1 is single bonded to O H, C 5 is single bonded to H at an angle farther from the viewer, and C 6 is single bonded to O H at angle farther from the viewer. In the fourth ring, C 1 is double bonded to O, C 5 is single bonded to O H and C 6 is single bonded to a C that is double bonded to O and single bonded to an N H subscript 2 group. In Tetracycline’s membrane soluble form, C 4 of the fourth ring is single bonded to a nitrogen that is single bonded to two C H subscript 3 groups. In Tetracycline’s water soluble form, C 4 of the fourth ring is single bonded to a positively charged nitrogen that is single bonded to two C H subscript 3 groups and single bonded to a hydrogen.
FIGURE 3.11 ■Common drugs are membrane-permeant weak acids and bases.A. The protonated form of the drug (HA) is hydrophobic and penetrates the cell membrane. The deprotonated charged form (A⁻) is soluble in the cytoplasm and in the bloodstream for drug delivery. B. The uncharged weak base (B) is hydrophobic and penetrates the membrane, whereas the protonated form (BH⁺) is soluble in the bloodstream.
Membrane-permeant weak acids and weak bases cross the membrane in their uncharged form: HA (weak acid) or B (weak base). On the other side, entering the aqueous cytoplasm, the acid dissociates (HA to A− and H+) or the base reassociates with H+ (B to BH+). In effect, membrane-permeant acids conduct acid (H+) across the membrane, causing acid stress; similarly, membrane-permeant bases conduct OH− across the membrane, causing alkali stress. If the H+ concentration (acidity) outside the cell is greater than inside, it will drive weak acids into the cell.
Many key substances in cell metabolism are membrane-permeant weak acids and weak bases, such as acetic acid. Most pharmaceutical drugs—therapeutic agents delivered to our tissues via the bloodstream—are weak acids or weak bases whose uncharged forms exist at sufficiently low concentration to cross the membrane without disrupting it. Examples of weak acids that deprotonate (acquiring negative charge) at neutral pH include aspirin (acetylsalicylic acid) and penicillin (Fig. 3.11A). Examples of weak bases that protonate (acquiring positive charge) at neutral pH include Prozac (fluoxetine) and tetracycline (Fig. 3.11B).
Thought Question
3.3 Amino acids have acidic and basic groups that can dissociate. Why are they not membrane-permeant weak acids or weak bases? Why do they fail to cross the phospholipid bilayer?
ANSWER ANSWER
At neutral pH, an amino acid has both a positively charged amine and a negatively charged carboxylate; that is, it can act as either a weak acid or a weak base. Charged ions will not freely pass through a plasma membrane (unless the molecule has an extremely hydrophobic group). In an amino acid, if either charged group becomes neutralized by acid or base, the other group remains charged, so the molecule as a whole will never cross the membrane.
Transmembrane ion gradients. Molecules that carry a fixed charge, such as hydrogen and sodium ions (H+ and Na+), cannot cross the phospholipid bilayer. Such ions usually exist in very different concentrations inside and outside the cell. An ion gradient (ratio of ion concentrations) across the cell membrane can store energy for nutrition or to drive the transport of other molecules. Inorganic ions require transport through specific transport proteins, or transporters. So, too, do organic molecules that carry a charge at cytoplasmic pH, such as amino acids and vitamins.
Transport may be passive or active. In passive transport, molecules accumulate or dissipate along their concentration gradient. Active transport—that is, transport from lower to higher concentration—requires cells to spend energy. A transport protein obtains energy for active transport by cotransport of another substance down its gradient from higher to lower concentration or by coupling transport to a chemical reaction.
Note: Active transport by proteins is covered in detail in Chapter 4.
To Summarize
The cell membrane is a bilayer of phospholipids. Microbes may change the composition of these lipids in response to their environment.
Archaeal membranes have ether-linked terpenoids, which confer increased stability at high temperature and extreme acidity. Some archaea have diglycerol tetraethers, which form a monolayer.
The membrane contains embedded proteins that have a hydrophobic surface. Membrane proteins serve diverse functions, including transport, cell defense, and cell communication.
Small uncharged molecules, such as oxygen, can penetrate the cell membrane by diffusion.
Membrane-permeant weak acids and weak bases exist partly in an uncharged form that can diffuse across the membrane and increase or decrease, respectively, the H+ concentration within the cell.
Ion gradients generated by membrane pumps store energy.
The major component of membranes. A typical phospholipid is composed of a core of glycerol to which two fatty acids and a modified phosphate group are condensed.
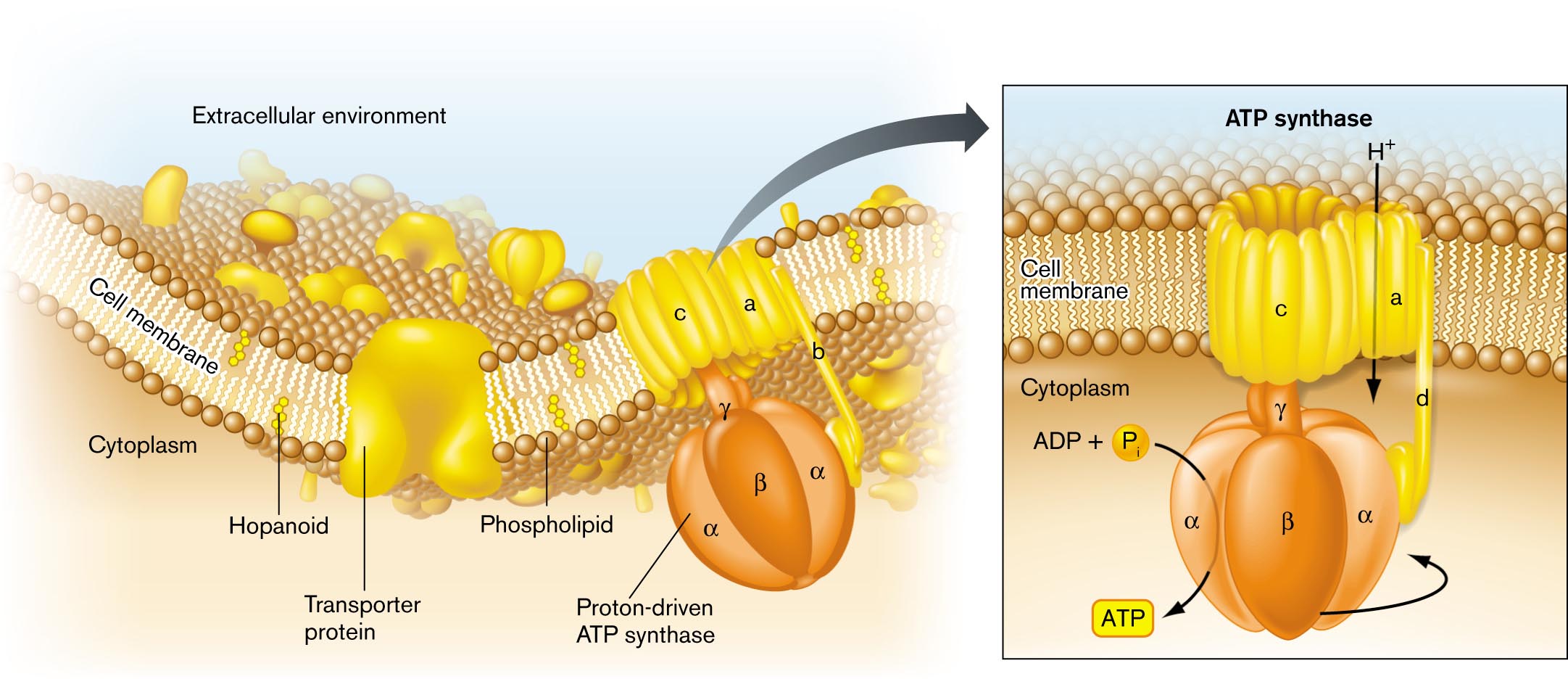
An illustration of the general structure of a bacterial cell membrane. The area beyond the cell membrane is labeled the extracellular environment. The area within the cell is labeled cytoplasm. The cell membrane is comprised of a phospholipid bilayer. Several hopanoids are layered within the membrane. A transporter protein embedded in the membrane opens to both the extracellular environment and the intracellular environment. A proton-driven A T P synthase is embedded in the membrane and opens to the both the extracellular environment and the intracellular environment. Further detail on the A T P synthase describes that hydrogen is taken into the cell. The proton driven pump is located in the cell interior, and uses A D P plus inorganic phosphate to generate A T P.
FIGURE 3.2 ■Bacterial cell membrane. The cell membrane consists of a phospholipid bilayer, in which the hydrophobic fatty acid chains are directed inward, away from water. The bilayer contains stiffening agents such as hopanoids. Half the membrane volume consists of proteins such as the ATP synthase protein complex.
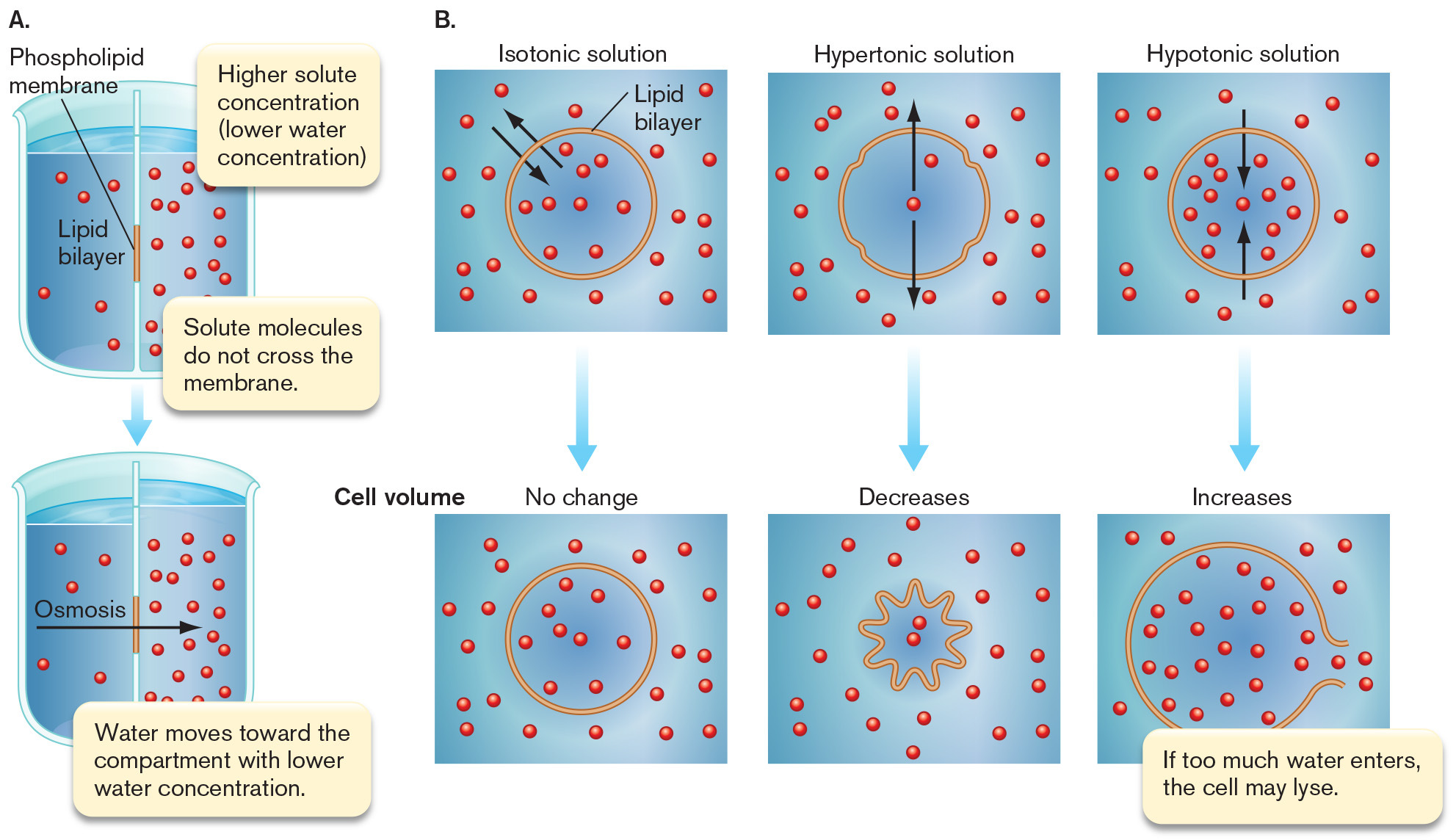
Two diagrams A and B depict osmosis and water balance in a cell. Part A shows two beakers split vertically down the center with water filling up either side. The divider down the center of each beaker represents the lipid bilayer. A portion in the center is highlighted and labeled Phospholipid membrane. The water contains small red dots representing a solute. In the top beaker, the right side has a higher solute concentration and a lower water concentration than the left side. The top inset reads higher solute concentration open parenthesis lower water concentration close parenthesis. The bottom inset reads solute molecules do not cross the membrane. An arrow points downward from the top beaker to the bottom beaker. The bottom beaker has less water and less solute on the left than the right side has. An arrow points from the left side to the right and is labeled Osmosis. Text reads water movies toward the compartment with lower water concentration. Part B is a three part diagram starting with an isotonic solution. The lipid bilayer is represented as a yellow circle and red molecules are scattered through the water and the cell. Two black arrows point into and out of the cell. A blue arrow points downward from here to a similar cell, that has experienced no change in volume. The second part of the diagram has a hypertonic solution. The lipid bilayer is represented as a yellow circle with four small indentations. There are two red molecules in the cell. Two black arrows point from inside of the cell to the outside. A blue arrow points downward from here to a star shaped cell. The volume of this cell has decreased and the two red molecules are closer together. The final part shows a hypotonic solution. The lipid bilayer is represented as a yellow circle. There are a lot of red molecules within the cell. Two black arrows point from outside of the cell to the inside. A blue arrow points downward from here to a larger cell. The cell volume has increased and the red molecules within it are all spread out. There’s an opening on the right side of the cell from which molecules are escaping. Text reads if too much water enters, the cell may lyse.
A diagram of two beakers split vertically down the center with water filling up either side. The divider down the center of each beaker represents the lipid bilayer. A portion in the center is highlighted and labeled Phospholipid membrane. The water contains small red dots representing a solute. In the top beaker, the right side has a higher solute concentration and a lower water concentration than the left side. The top inset reads higher solute concentration open parenthesis lower water concentration close parenthesis. The bottom inset reads solute molecules do not cross the membrane. An arrow points downward from the top beaker to the bottom beaker. The bottom beaker has less water and less solute on the left than the right side has. An arrow points from the left side to the right and is labeled Osmosis. Text reads water movies toward the compartment with lower water concentration.
A three part diagram starting with an isotonic solution. The lipid bilayer is represented as a yellow circle and red molecules are scattered through the water and the cell. Two black arrows point into and out of the cell. A blue arrow points downward from here to a similar cell, that has experienced no change in volume. The second part of the diagram has a hypertonic solution. The lipid bilayer is represented as a yellow circle with four small indentations. There are two red molecules in the cell. Two black arrows point from inside of the cell to the outside. A blue arrow points downward from here to a star shaped cell. The volume of this cell has decreased and the two red molecules are closer together. The final part shows a hypotonic solution. The lipid bilayer is represented as a yellow circle. There are a lot of red molecules within the cell. Two black arrows point from outside of the cell to the inside. A blue arrow points downward from here to a larger cell. The cell volume has increased and the red molecules within it are all spread out. There’s an opening on the right side of the cell from which molecules are escaping. Text reads if too much water enters, the cell may lyse.
FIGURE A2.5 ■Osmosis and water balance.A. Osmosis. B. Movement of water across the cell membrane, and shrinkage or expansion of the membrane in isotonic, hypertonic, and hypotonic environments. Black arrows indicate net water movement.
 ANSWER
ANSWER ANSWER
ANSWER