How do bacteria and archaea protect their cell membrane from a dangerous environment? For most species, the cell envelope includes at least one structural supporting layer, like an external skeleton, located outside the cell membrane. As seen in Figure 3.1, the most common structural support is the cell wall (Fig. 3.12). Many species possess additional coverings, such as an outer membrane or an S-layer. Nevertheless, a few prokaryotes, such as the mycoplasmas, have a cell membrane with no outer layers and depend on host fluids for osmotic balance. Some archaea with only a cell membrane grow in extreme acid (pH zero); for example, Ferroplasma, found in iron mines. How their cells survive is unknown.
More information
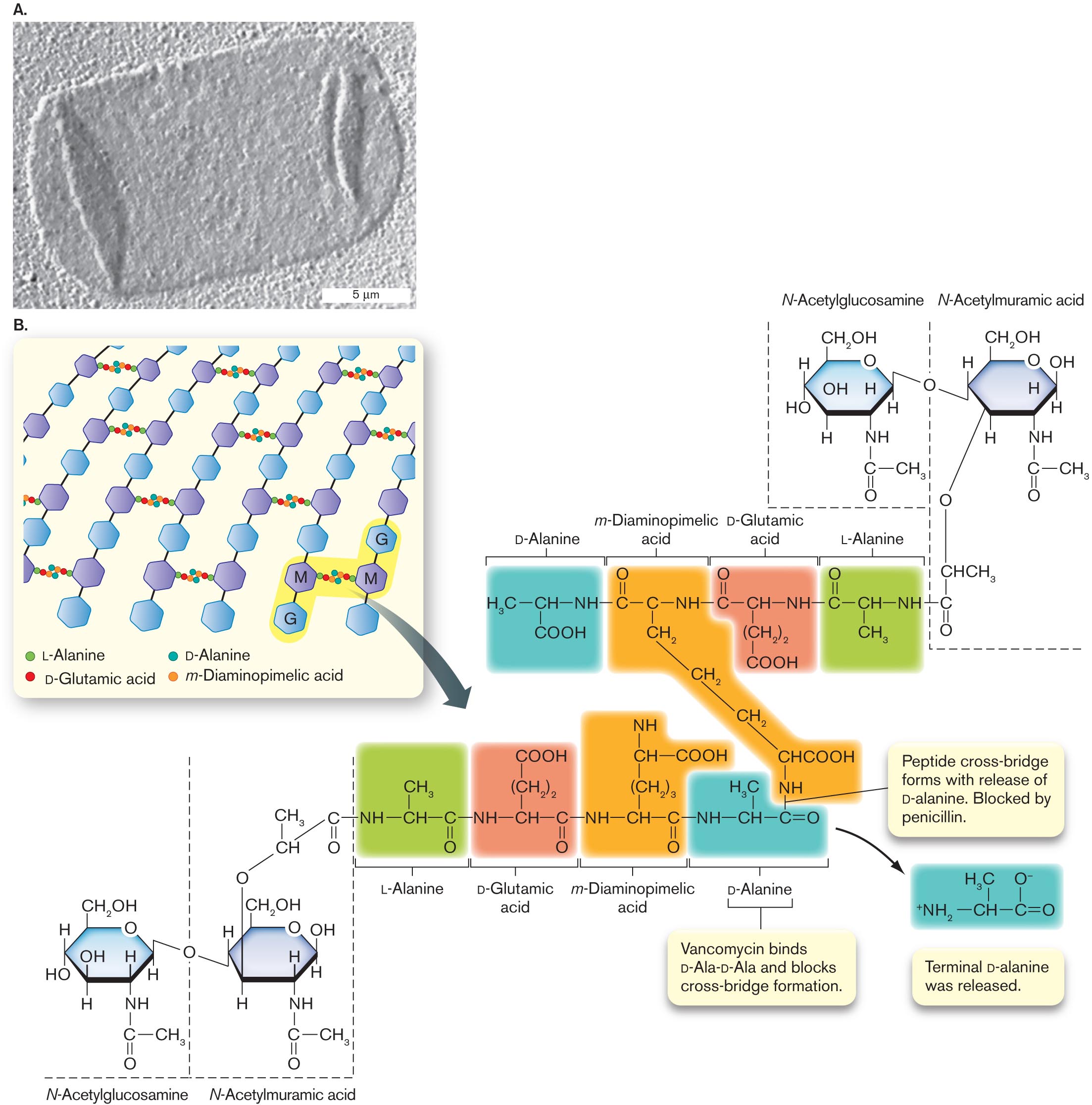
A transmission electron micrograph of an isolated sacculus and an illustration of peptidoglycan structure.
A transmission electron micrograph of an isolated sacculus from Escherichia coli. The sacculus is a rectangular shape of about 20 micrometers in length and 10 micrometers in width. The sacculus appears transparent in the micrograph.
An illustration of peptidoglycan structure. It consists of several benzenes bonded vertically labeled as G and M alternatively. The text at the bottom reads L-Alanine, D-Glutamic acid, D-Alanine, and m-Diaminopimelic acid. G and M are shaded and is bonded with the alanine and acids. An arrow pointing toward the molecular structures of L-Alanine, D-Glutamic, D-Alanine acid, and m-Diaminopimelic acid. The structure of D-Alanine consists of C H in the center is single bonded to N H at the right, H 3 C at the left, and C O O H at the bottom. N H is single bonded to the structure of m-Diaminopimelic acid that consists of carbon in the center is single bonded to N H at the right, single bonded to carbon at the left, and single bonded to methyl at the bottom. The carbon in the left is double-bonded to oxygen at the top. Methyl at the bottom is single-bonded two methyl groups at the bottom and single bonded to C H C O O H is single bonded to N H. N H at the right is single bonded to the structure of D-Glutamic acid that consists of C H in the center is single bonded to N H at the right, carbon at the left, and (C H 2) 2 at the bottom and is single bonded to C O O H at the bottom. N H at the right is single bonded to the structure of L-Alanine that consists of C H in the center is single bonded to N H at the right, carbon at the left, and single bonded to methyl at the bottom. The carbon at the left is double-bonded to oxygen. Therefore the structures of L-Alanine, D-Glutamic, D-Alanine acid, and m-Diaminopimelic acid at the bottom are highlighted differently. N H at the bottom of the structure m-Diaminopimelic Acid is single bonded to carbon at the right in D-Alanine at the bottom. A bond between N H and Carbon reads Peptide cross-bridge forms with the release of D-alanine. They are blocked by penicillin. The text pointing toward the structure of D-Alanine reads Vancomycin binds D-Ala-D-Ala and blocks cross-bridge formation. An arrow from the D-Alanine structure leads to the structure that consists of C H in the center is single bonded to carbon in the right, single bonded to ammonia at the left, and double-bonded to oxygen at the right. C H in the center is single bonded to H 3 C at the top, and carbon at the right is single bonded to oxygen with minus ion at the top. Two structures of N-Acetylglucosamine and N-Acetylmuramic acid are bonded to the L-Alanine structure at the top right, and L-Alanine structure at the bottom left. The N-Acetylglucosamine structure consists of benzene in which C 1 is single bonded to C H 2 O H, C 2 is substituted by oxygen, C 3 is single bonded to hydrogen, C 4 is single bonded to N H at the bottom is single bonded to carbon at the bottom, in which carbon is single bonded to methyl at the right and double-bonded to oxygen at the bottom. C 5 is single bonded to hydrogen at the bottom, and C 6 is single bonded to hydroxyl and hydrogen. The N-acetylmuramic acid consists of benzene in which C 1 is single bonded to C H 2 O H, C 2 is substituted by oxygen, C 3 is single bonded to hydrogen, C 4 is single bonded to N H at the bottom is single bonded to carbon at the bottom, in which carbon is single bonded to methyl at the right and double-bonded to oxygen at the bottom. C 5 is single bonded to hydrogen at the bottom, and C 6 is single bonded to hydroxyl and hydrogen, in which hydrogens in C 6 of the two structures ate bonded to oxygen. C 5 of the N-Acetylmuramic acid is single bonded to oxygen at the bottom is single bonded to C H C H 3, which is single bonded to carbon and the carbon is double-bonded to oxygen at the bottom, and the carbon is single bonded to L- Alanine structure of N H at the right.
FIGURE 3.12 ■The cell wall: peptidoglycan sugar chains and cross-bridges.A. Isolated sacculus (entire cell wall) from Escherichia coli (TEM). B. A disaccharide unit of glycan has an attached peptide of four to six amino acids. W. VOLLMER ET AL. 2008. FEMS MICROBIOL. REV.32:149, FIG. 3A
The Cell Wall Is a Single Molecule
The bacterial cell wall, also known as the sacculus, consists of a single interlinked molecule that envelops the cell. The sacculus has been isolated from Escherichia coli and visualized by transmission electron microscopy (TEM); in Figure 3.12A, the isolated sacculus appears flattened on the sample grid like a deflated balloon. Its geometrical structure encloses maximal volume with minimal surface area. The sacculus—unlike the membrane—is a single-molecule cage-like structure, highly porous to ions and organic molecules. The mesh grows by strand insertion and elongation in arcs around the cell. The cage-like form is not rigid; it is more like a flexible mesh bag with unbreakable joints. Turgor pressure within the enclosed cytoplasm fills out the cell’s shape, whether elongated rod, spherical coccus, or other.
Peptidoglycan structure. Most bacterial cell walls are composed of peptidoglycan, a polymer of peptide-linked chains of amino sugars. Peptidoglycan is synonymous with murein (“wall molecule”). The molecule consists of parallel polymers of disaccharides called glycan chains cross-linked with peptides of four to six amino acids (Fig. 3.12B). Peptidoglycan is unique to bacteria, although some archaea build analogous structures whose overall physical nature is similar. (Archaeal cell walls are presented in Chapter 19.)
The long chains of peptidoglycan consist of repeating units of a disaccharide composed of N-acetylglucosamine (an amino sugar derivative) and N-acetylmuramic acid (glucosamine plus a lactic acid group; Fig. 3.12B). The lactate group of muramic acid forms an amide link with the amino terminus of a short peptide containing four to six amino acid residues. The peptide extension can form cross-bridges connecting parallel strands of glycan.
The peptide contains two amino acids in the unusual d mirror form: d-glutamic acid and d-alanine. The third amino acid, m-diaminopimelic acid, has an extra amine group, which forms an amide link to a cross-bridged peptide. The amide link forms with the fourth amino acid of the adjacent peptide, d-alanine (Fig. 3.12B). Removal of a second d-alanine at the end of the chain forms the cross-bridge. The cross-linked peptides of neighboring glycan strands form the cage of the sacculus.
Note: Amino acids have two forms that are mirror opposites, d and l, of which only the l form is incorporated by ribosomes into protein. The d-form amino acids, however, are used by microbes for many nonprotein structural molecules.
The details of peptidoglycan structure vary among bacterial species. Some Gram-positive species, such as Staphylococcus aureus (a cause of hospital-acquired infections), have peptides linked by bridges of pentaglycine instead of the d-alanine link to m-diaminopimelic acid. In Gram-negative species, the m-diaminopimelic acid is linked to the outer membrane, as will be discussed shortly.
Peptidoglycan synthesis as a target for antibiotics. Synthesis of peptidoglycan requires many genes encoding enzymes to make the special sugars, build the peptides, and seal the cross-bridges (see Fig. 3.12B). Many of these enzymes bind the antibiotic penicillin and are thus known as penicillin-binding proteins. Because peptidoglycan is unique to bacteria, enzymes of peptidoglycan biosynthesis make excellent targets for new antibiotics. For example, the transpeptidase that cross-links the peptides is the target of penicillin. Vancomycin, a major defense against Clostridioides difficile and drug-resistant staphylococci, prevents cross-bridge formation by binding the terminal d-Ala-d-Ala dipeptide. Vancomycin binding prevents release of the terminal d-alanine.
Unfortunately, the widespread use of such antibiotics selects for evolution of resistant strains. One of the most common agents of resistance is the enzyme beta-lactamase, which cleaves the lactam ring of penicillin, rendering it ineffective as an inhibitor of transpeptidase (also called penicillin-binding protein 2). In a different mechanism, strains resistant to vancomycin contain an altered enzyme that adds lactic acid to the end of the branch peptides in place of the terminal d-alanine. The altered peptide is no longer blocked by vancomycin. As new forms of drug resistance emerge, researchers continue to seek new antibiotics that target cell wall formation (discussed in Chapter 27).
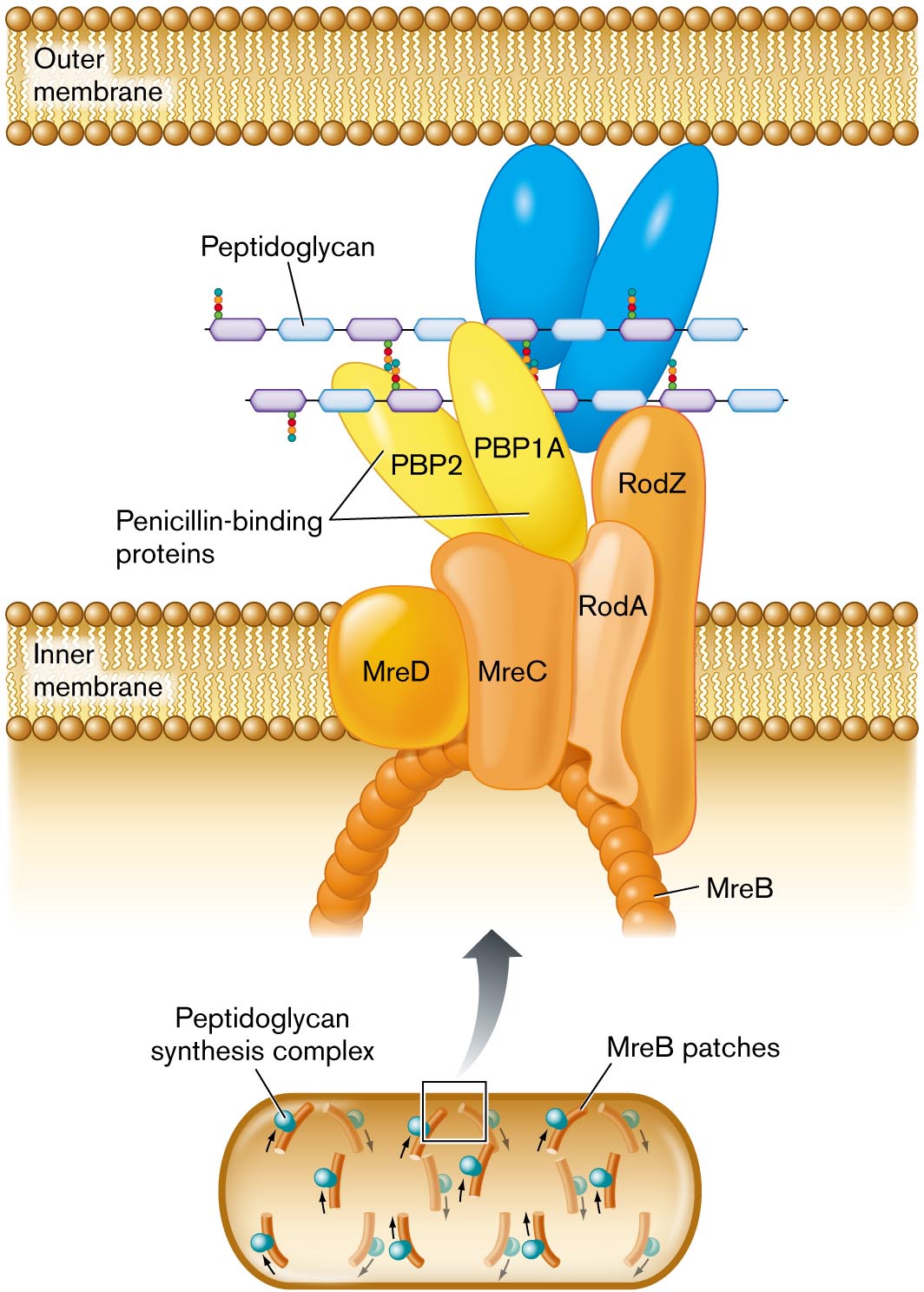
How does peptidoglycan grow, overall, throughout the elongating cell? A peptidoglycan synthesis complex extends the chains of amino-sugars. So-called penicillin-binding proteins catalyze the formation of peptide cross-bridges (Fig. 3.13). These proteins were named for their property of binding the antibiotic penicillin, which disables cross-bridge formation (details in Chapter 27). The overall direction of cell wall extension is organized by a protein complex that includes MreB. MreB polymerizes in a helical direction along an arc beneath the plasma membrane (inner membrane, for a Gram-negative bacterium). The RodA and RodZ proteins are needed to shape the cell as a rod (bacillus).
More information
A detailed illustration of peptidoglycan synthesis. Synthesis is shown to take place between the outer membrane and the inner membrane. Synthesis is mediated by a peptidoglycan synthesis complex, shown blown-up in the illustration. The complex extends from the interior of the cell across the inner membrane into the space between the inner and outer membranes. An arch shaped structure supports the complex, the structure is comprised of M r e B patches. Attached to the arch are several irregularly shaped proteins labeled M r e D, M r e C, R o d A, and R o d Z. Two penicillin binding proteins sit atop M r e C. The penicillin binding proteins are labeled P B P 2 and P B P 1 A. A chain of peptidoglycan is synthesized through the complex.
FIGURE 3.13 ■Peptidoglycan synthesis is organized by penicillin-binding proteins (PBP2, PBP1A) and by cytoskeletal proteins. Protein MreB guides the direction of synthesis in helical arcs around the cell.
Thought Question
3.4 What genetic experiments could you propose to test the model of envelope expansion shown in Figure 3.13?
ANSWER ANSWER
Use mutagenesis (treatment with a mutagen) to generate E. coli strains containing point mutations in rodA or rodZ. Observe growth of the mutant cells. Prediction: Some of the rodA or rodZ mutants will have defective RodA or RodZ proteins that cause inefficient cell wall synthesis. The mutant cells grow slowly, as bulging, blob-shaped cells, not as rods. Now test the question, How does RodA or RodZ interact with other components of the envelope extension complex? Introduce new mutations in genes encoding proteins of the peptidoglycan extension complex (Fig. 3.13), such as mreB and pbp2. With the newly introduced mutations, we find that some of the rodZ mutants revert to normal rod-shaped growth. Thus, altered MreB or Pbp2 proteins might compensate for the alteration in RodZ. This result suggests that the RodZ protein could interact with MreB and Pbp2 in the extension complex. For research published on these questions, see Daisuke Shiomi et al. 2013. Mol. Microbiol.87:1029.
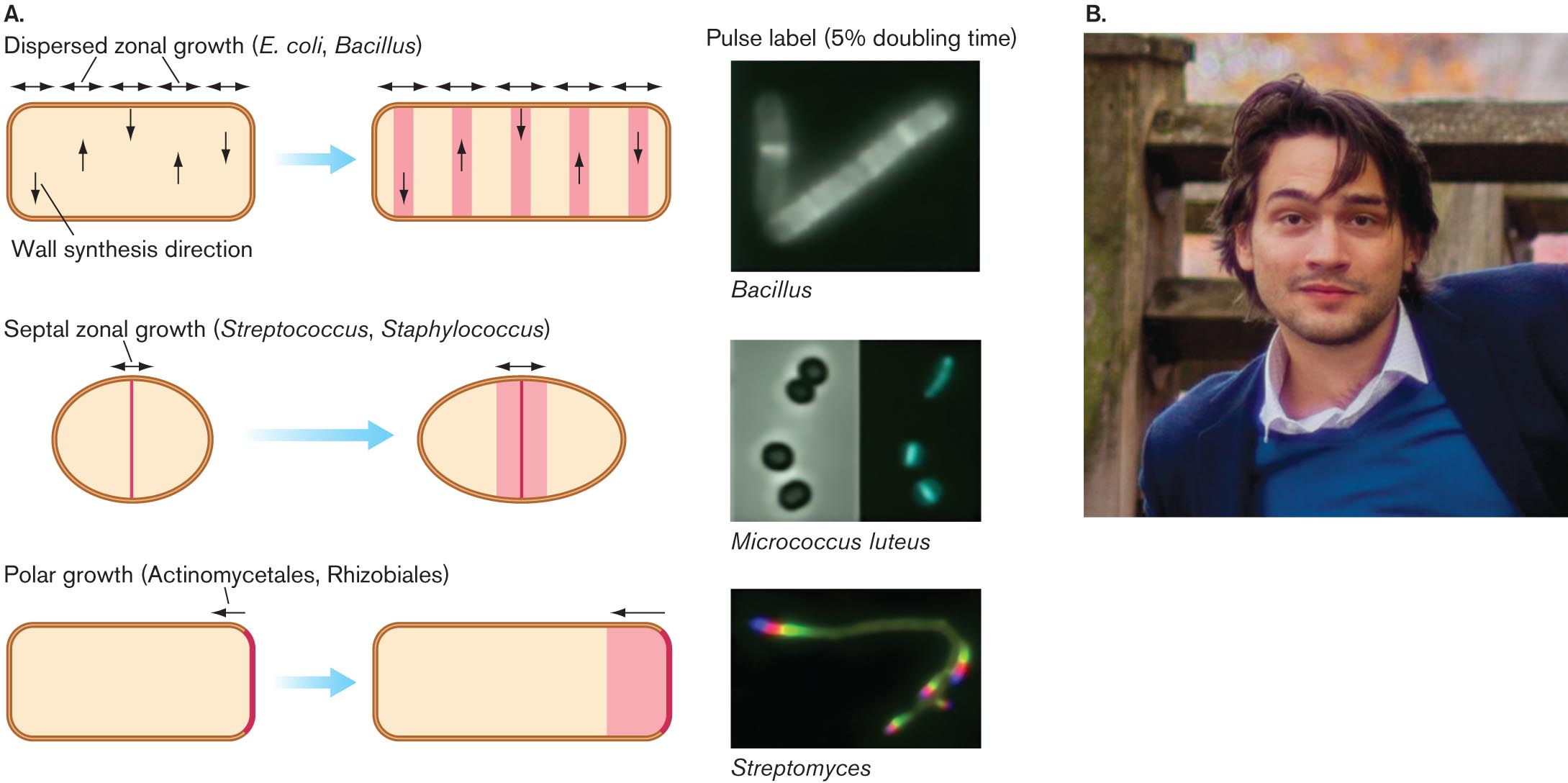
Notably, different kinds of bacteria have evolved to organize their cell wall growth differently. Yves Brun and his student Erkin Kuru, at Indiana University, devised an ingenious way to reveal the growth pattern (Fig. 3.14). Kuru designed fluorophore-tagged d-amino acids that the growing cell wall incorporates (such as d-alanine; see Fig. 3.12). Because these are d-amino acids and not l-amino acids, ribosome-directed translation does not use them. Thus, the fluorophore-linked d-amino acids label cell wall only, not proteins.
More information
An illustration of peptidoglycan growth in different species and a photo of Erkin Kuru.
An illustration shows variations in peptidoglycan growth between different bacterial species. The first growth type is dispersed zonal growth, with E. coli or Bacillus species as examples. A rod shaped cell is shown. Horizontal double-headed arrows are along the top wall and represent dispersed zonal growth of the peptidoglycan. Up and down arrows inside of the cell represent wall synthesis direction. The cell is shown again, with peptidoglycan growing in the direction of the arrows. An accompanying micrograph shows the pulse label, or 5 percent doubling time, of a Bacillus bacterium. Fluorescent zones are visible within the cell when they are shown in the illustration. The second growth type is spetal zonal growth, with Streptococcus or Staphylococcus as examples. An oval-shaped cell is shown. A vertical line runs down the center. A double-headed arrow at the top represents septal zonal growth of the peptidoglycan. The cell is shown again with peptidoglycan growing in the direction of the arrows. An accompanying micrograph shows the pulse label for Micrococcus luteus with four coccoid cells. The micrograph is shown again with thin fluorescent zones in the center of the cells. The third growth type is polar growth, with Actinomycetales and Rhizobiales as examples. A rod shaped cell is shown. The right edge of the cell is highlighted, and an arrow points to the left showing polar growth. The cell is shown again with a large section of peptidoglycan growing in the direction of the arrow. An accompanying micrograph shows the pulse label for Streptomyces. Fluorescent zones are visible at both ends of the long filamentous cell.
Micrographs of pulse labeling in Bacillus, Micrococcus luteus, and Streptomyces. In the micrograph of Bacillus, two rod shaped bacteria are seen. Within the bacteria, multiple fluorescent bands are visible. In the micrograph of Micrococcus luteus, four coccoid bacteria. Within the bacteria, single, thin, fluorescent bands are visible at their centers. In the micrograph of Streptomyces, a long filamentous bacterium is seen. Within the bacterium, fluorescent zones are visible at each end of the cell.
A photo of Erkin Kuru. He has brown hair, brown eyes, and some facial hair. He is smiling at the camera in the photo.
Kuru found that bacteria such as E. coli and Bacillus subtilis synthesize cell wall in zones dispersed throughout the cell. Gram-positive cocci, however, such as Streptococcus and Staphylococcus, synthesize cell wall only at the midpoint (or septum), where the cell is about to fission (discussed in Section 3.4). Still other bacteria, such as the actinomycete Streptomyces, form new cell wall only at the poles. Cell growth is discussed further in Section 3.4.
Thought Question
3.5 What other ways can you imagine that bacteria might mutate to become resistant to vancomycin?
ANSWER ANSWER
A common means of resistance to antibiotics is to pump them out of the cell. A protein pump that exports other molecules might mutate to capture vancomycin and export it from the cell. Another possibility is that an enzyme could modify the vancomycin by adding phosphoryl groups or acetyl groups, which would prevent the antibiotic from binding the alanine dipeptide. Still another possibility is that the bacteria might evolve a thicker cell wall that would exclude the vancomycin from the inner layers of peptidoglycan.
Cell Envelope of Bacteria
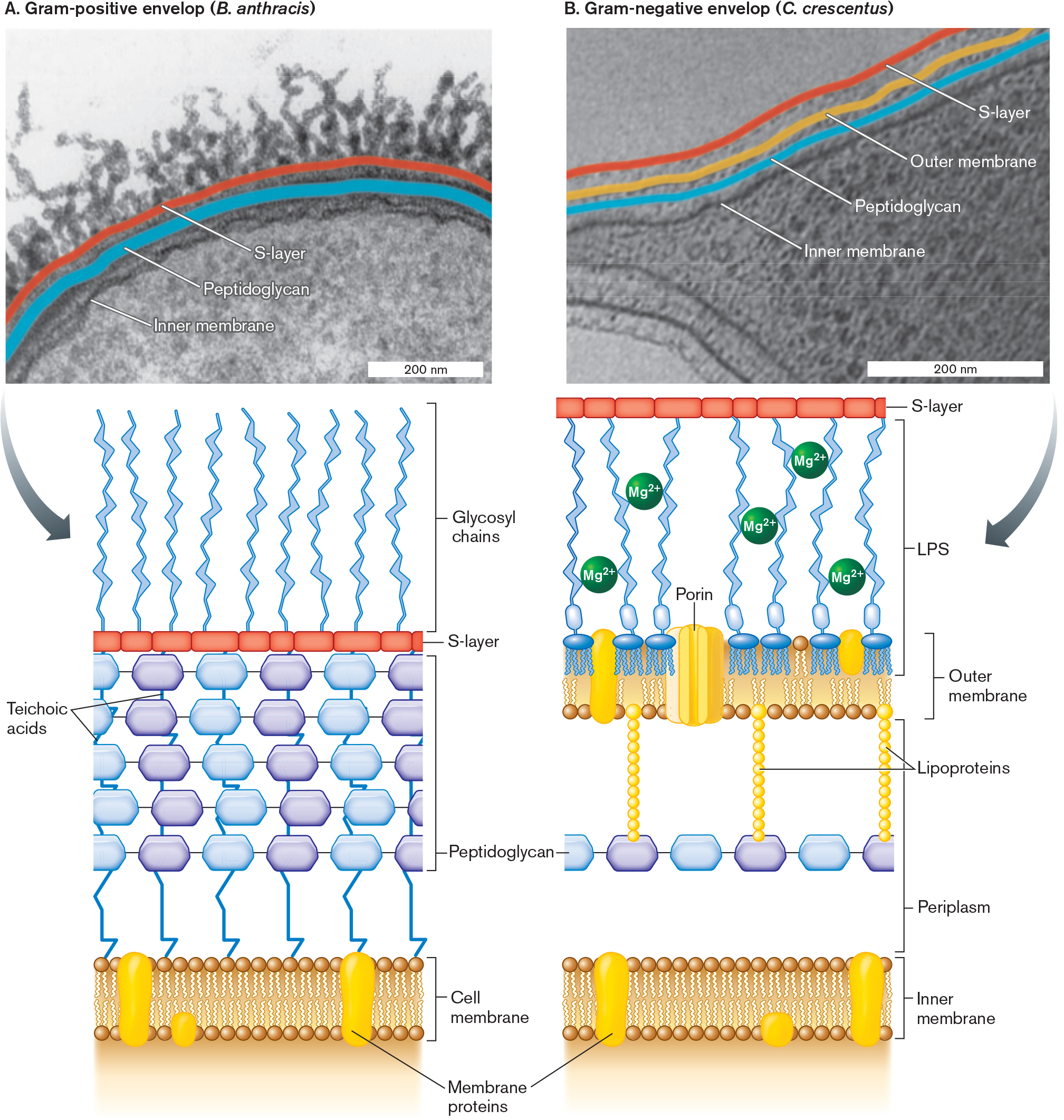
Most bacteria have additional envelope layers that provide structural support and protection from predators and host defenses (Fig. 3.15). Additional molecules are attached to the cell wall and cell membrane, and some thread through the layers. Here we present the envelope composition of three major kinds of bacteria, two of which (Firmicutes and Proteobacteria) are distinguished by the Gram stain. The third, Mycobacteria, is distinguished by the acid-fast stain.
More information
Two micrographs of bacterial cell envelopes with accompanying illustrations of envelope structure are shown.
A micrograph of a Gram-positive cell envelope, with structures labeled. Bacillus anthracis is the bacteria in the example. The micrograph is magnified so that only a portion of the cell is visible. The cell appears as a curving half-circle with tendrils lining the cell’s outer edge. The visible part of the cell is about 600 nanometers wide and 400 nanometers long. The tendrils are each about 200 nanometers long. The inner membrane is identified. Exterior to the inner membrane is the peptidoglycan layer, and beyond that is the S-layer. The peptidoglycan layer and the S layer are identified as components of the cell envelope. An illustration beneath the micrograph describes the cell envelope structure in greater detail.
A detailed illustration of the structure of a Gram-positive cell envelope. The cell membrane consists of a phospholipid bilayer with several membrane proteins embedded. Exterior to the membrane is a thick layer of peptidoglycan, connected by teichoic acids. Beyond the peptidoglycan layer is a thin S layer. Beyond the S layer are thin tendrils identified as glycosyl chains.
A micrograph of a Gram-negative cell envelope, with structures labeled. Caulobacter crescentus is the bacteria in the example. The cell is irregularly shaped and only partly visible. The visible portion is about 600 nanometers long and 400 nanometers wide at its widest point. The cell is tapered at one end. The inner membrane is identified. Exterior to the inner membrane is a peptidoglycan layer, followed by the outer membrane, and then an S layer. The peptidoglycan layer, the outer membrane, and the S-layer are identified as components of the cell envelope.
A detailed illustration of the structure of a Gram-negative cell envelope. The inner membrane consists of a phospholipid bilayer with membrane proteins embedded. Beyond the inner membrane, a thin layer of peptidoglycan is suspended in periplasm. The peptidoglycan layer is connected to the outer membrane by chains of lipoproteins. The outer membrane is lined with phospholipids on the side facing the peptidoglycan. A porin larger than the membrane is embedded in the outer membrane. The other side of the outer membrane is part of the L P S layer. The L P S layer consists of glycosyl chains and M g superscript 3 plus molecules. The S-layer is a thin layer beyond the L P S.
FIGURE 3.15 ■Cell envelope: Gram-positive (Bacillus anthracis) and Gram-negative (Caulobacter crescentus).A. Firmicutes (Gram-positive) cells have a thick cell wall with multiple layers of peptidoglycan, threaded by teichoic acids. The micrograph shows the Gram-positive envelope of Bacillus anthracis (TEM). B. Proteobacteria (Gram-negative) cells have a single layer of peptidoglycan covered by an outer membrane; the cell membrane is called the inner membrane. The outer membrane lipopolysaccharides (LPS) are cross-linked by positively charged magnesium ions (Mg2⁺). The micrograph shows the Gram-negative envelope of Caulobacter crescentus (TEM). LPS = lipopolysaccharides. S. MESNAGE ET AL. 1998. J BACTERIOL.180:52–58T. BHARAT ET AL. 2017. NAT MICROBIOL.2:17059
Firmicutes (Gram-positive) have a thick cell wall with 3–20 layers of peptidoglycan, interpenetrated by teichoic acids (Fig. 3.15A). The phylum Firmicutes consists of Gram-positive species such as Bacillus thuringiensis and Streptococcus pyogenes, the cause of strep throat.
Proteobacteria (Gram-negative) have a thin cell wall with one or two layers of peptidoglycan, enclosed by an outer membrane (Fig. 3.15B). Included among the phylum Proteobacteria are Gram-negative species such as Escherichia coli and nitrogen-fixing Sinorhizobium meliloti.
Mycobacteria of the phylum Actinomycetes have a complex, multilayered envelope that includes defensive structures such as mycolic acids. Examples include Mycobacterium tuberculosis (the cause of tuberculosis) and M. leprae (the cause of leprosy).
Note that other important kinds of bacteria, such as cyanobacteria (see Fig. 18.8) and spirochetes (see Fig. 18.43), have very different envelope structures. These different envelopes may stain Gram-positive, Gram-negative, or variable (discussed in Chapter 18). Archaea have yet other diverse kinds of envelopes that cannot be distinguished by Gram stain (see Chapter 19).
Thought Question
3.6Figure 3.15 highlights the similarities and differences between the cell envelopes of Gram-negative and Grampositive bacteria. What do you think are the advantages and limitations of a cell having one layer of peptidoglycan (Gramnegative) versus several layers (Gram-positive)?
ANSWER ANSWER
Having multiple layers of peptidoglycan increases the cell’s resistance to osmotic shock, to desiccation stress, and to enzymes that cleave the cell wall. On the other hand, to build the layers of peptidoglycan requires more energy and biomass. In addition, a thick cell wall can slow the uptake of nutrients. The mycobacteria, which have exceptionally thick cell walls, grow very slowly.
Firmicute Cell Envelope—Gram-Positive
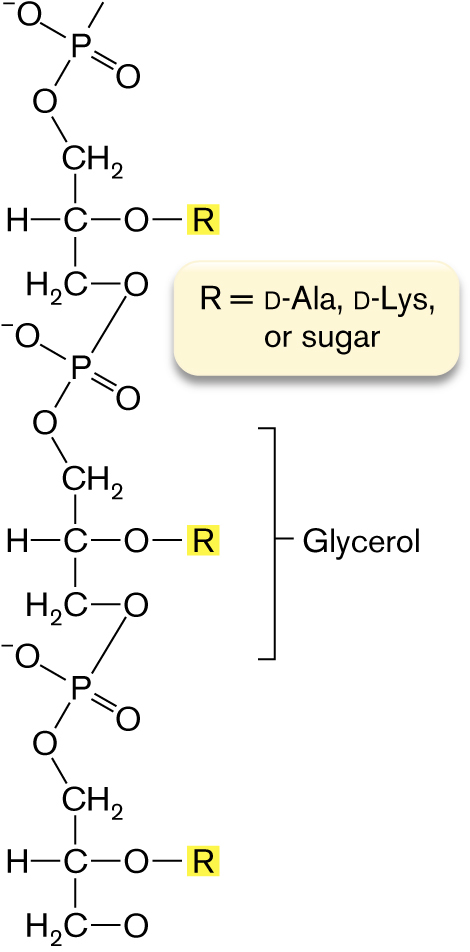
A section of a firmicute (Gram-positive) cell envelope is shown in Figure 3.15A. The multiple layers of peptidoglycan are reinforced by teichoic acids threaded through its multiple layers. Teichoic acids are chains of phosphodiester-linked glycerol or ribitol, with sugars or amino acids linked to the middle –OH groups (Fig. 3.16). The negatively charged cross-threads of teichoic acids, as well as the overall thickness of the Gram-positive cell wall, help retain the Gram stain.
More information
A molecular structure of Teichoic acid. The structure consists of three glycerol structures in which carbon is centered single bonded to methyl at the top, hydrogen at the left, oxygen at the right, and oxygen is single bonded to R is shown highlighted, and single bonded to H 2 C is single bonded to oxygen. The methyl at the top is single bonded to oxygen, in which oxygen is single bonded to phosphorous, and phosphorous is single bonded to oxygen with negative ion and double-bonded to oxygen. Oxygen at the bottom is bonded to the phosphate group at the bottom, in which oxygen is the group is bonded to the structure of glycerol. Further oxygen in the structure bonded to phosphate group and oxygen from the phosphate group is bonded to methyl of the glycerol structure at the bottom. A note reads R equals D-A l a, D-L y s, or sugar.
FIGURE 3.16 ■Teichoic acids. Teichoic acids in the Gram-positive cell wall consist of glycerol or ribitol phosphodiester chains.
How does the cell wall attach extracellular structures? Gram-positive bacteria have a type of enzyme called sortase that forms a peptide bond from a cell wall cross-bridge to a protein extending from the cell. Proteins attached by sortases can help the cell acquire nutrients or help the cell adhere to a substrate.
S-layer. Many free-living species of bacteria and archaea possess a tough surface layer called the S-layer. An S-layer is composed of protein subunits that fit together like tiles. This tough layer may provide defense against phages or predators. The S-layer is rigid, but it also flexes and allows substances to pass through it in either direction.
A Gram-positive example is Lysinibacillus sphaericus, a firmicute found on beets and carrots (Fig. 3.17A). Its S-layer is a crystalline sheet of thick subunits consisting of protein or glycoprotein (proteins with attached sugars) (Fig. 3.17B). Each subunit contains a pore large enough to admit a wide range of molecules. As modeled by cryo-EM tomography (see Chapter 2), the S-layer proteins are arranged in a highly ordered tetragonal array. A Gram-negative example is Caulobacter crescentus, a stalked proteobacterium found in ponds and wastewater (Fig. 3.17C). Its S-layer is packed in a hexagonal array (Fig. 3.17D). S-layers help pathogens such as Bacillus anthracis bind and attack host cells. An S-layer contributes to biofilm formation (the periodontal bacterium Tannerella forsythia) and swimming (the aquatic cyanobacterium Synechococcus species).
More information
Four high magnification micrographs of S-layer structure.
An image produced by atomic force microscopy of Lysinibacillus sphaericus. A few rod shaped cells are visible against a grainy background. The cells are between 4 to 6 micrometers long and 1 micrometer wide.
An image produced by atomic force microscopy of the S-layer surface. The surface is grid like, with square shaped units of about 10 nanometers in length and width.
An image produced by cryo- electron microscopy of Caulobacter crescentus, with S-layer units identified. A rod shaped cell is partly visible. The cell is 100 nanometers wide and greater than 600 nanometers long. Multi-colored dots follow a loose cylindrical shape around the cell. Overlaying text reads, hexamer positions plotted on tomogram.
A computational model of an S-layer unit from Caulobacter crescentus. The unit consists of a central hexagonal shape surrounded by 6 hexagonal shapes which are then surrounded by more hexagonal shapes. Each hexagonal unit has a side of about 11 nanometers in length.
FIGURE 3.17 ■S-layer reconstructions from microscopy.A.Lysinibacillus sphaericus imaged by atomic force microscopy (AFM). B. S-layer surface of L. sphaericus imaged by AFM. C.Caulobacter crescentus imaged by cryo-EM; S-layer subunits are marked. D.C. crescentus S-layer unit; computational model based on cryo-EM. TOBIAS GÜNTHER ET AL. 2014. RSC ADV.4 (93)TOBIAS GÜNTHER ET AL. 2014. RSC ADV.4 (93)T. BHARAT ET AL. 2017. NAT MICROBIOL.2:17059T. BHARAT ET AL. 2017. NAT MICROBIOL.2:17059
The functions of the S-layer are hard to study in the laboratory because the S-layer is often lost by bacteria after repeated subculturing. Traits commonly disappear in the absence of selective pressure for genes encoding them—a process called reductive evolution (discussed in Chapter 17). For example, the mycoplasmas are close relatives of Gram-positive bacteria that have permanently lost their cell walls, as well as the S-layer. Mycoplasmas have no need for cell walls, because they are parasites living in host environments, such as the human lung, where they are protected from osmotic shock.
Capsule. Another common extracellular structure is the capsule, a slippery layer of loosely bound polysaccharides. The capsule of pathogens such as Staphylococcus aureus can prevent phagocytosis by white blood cells, thereby enabling the pathogen to persist in the blood.
Thought Question
3.7 Why would laboratory culture conditions select for evolution of cells lacking an S-layer?
ANSWER ANSWER
Degeneration of protective traits is a common problem when conducting research on microbes that can produce 30 generations overnight. Their rapid reproductive rate gives ample opportunity for spontaneous mutations to accumulate over an experimental timescale. In the case of the S-layer, in a laboratory test tube free of predators or viruses, mutant bacteria that fail to produce the thick protein layer would save energy compared to S-layer synthesizers, and would therefore grow faster. Such mutants would quickly take over a rapidly growing population.
Proteobacterial Cell Envelope—Gram-Negative
The cell envelope of the Proteobacteria (Gram-negative) includes one or two layers of peptidoglycan covered by an outer membrane (Fig. 3.18). While free-living species such as C. crescentus have an S-layer, enteric bacteria such as E. coli often lack an S-layer. The Gram-negative outer membrane confers defensive abilities and toxigenic properties on many pathogens, such as Salmonella species and enterohemorrhagic E. coli (strains that cause hemorrhaging of the colon). Between the outer and inner (cell) membranes, the aqueous compartment (containing the cell wall) is called the periplasm.
More information
An illustration of the Gram-negative cell envelope structure and a micrograph of a murein lipoprotein mutant.
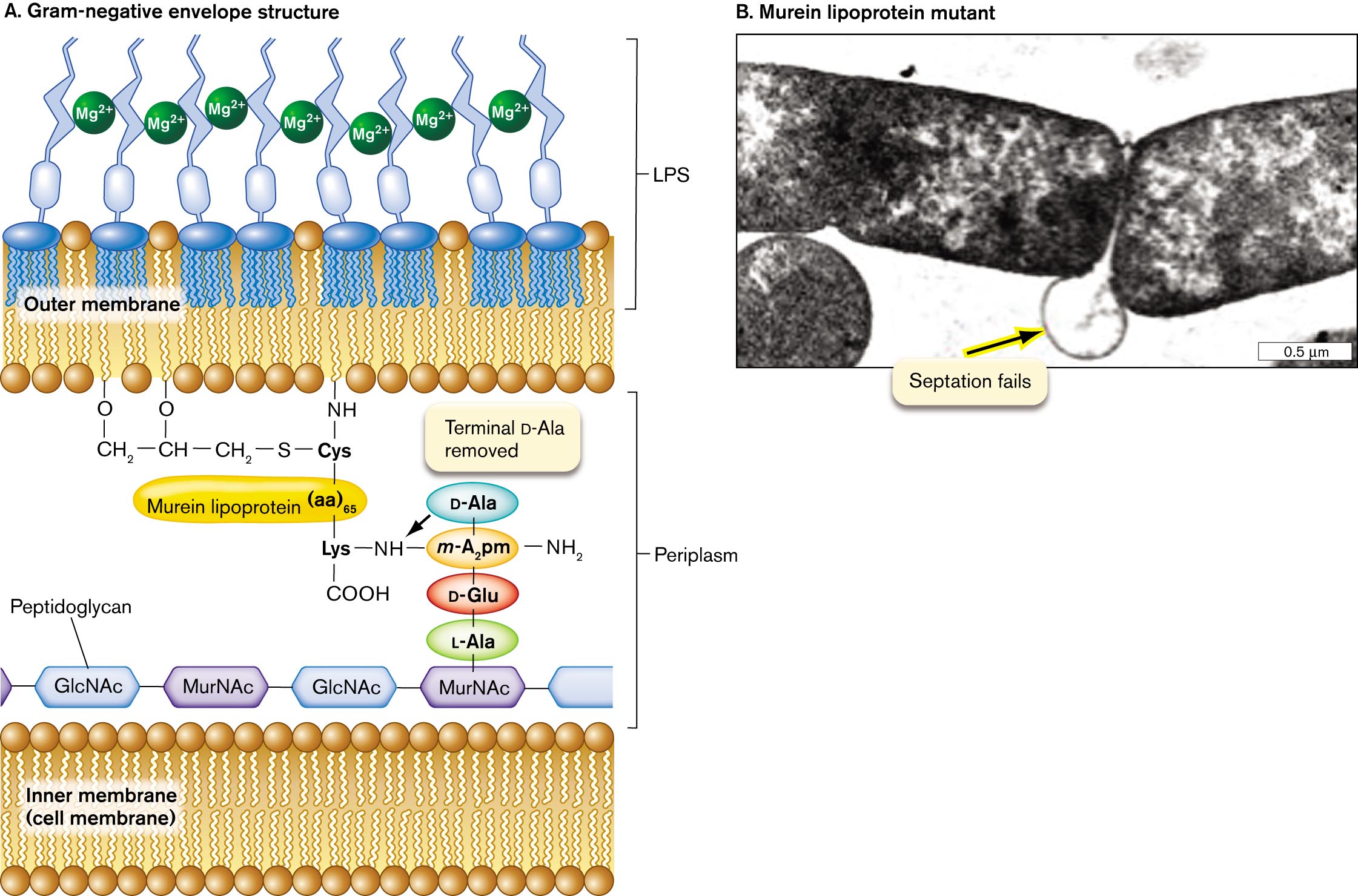
An illustration of the Gram-negative cell envelope structure. The inner cell membrane is shown as a phospholipid bilayer. Peptidoglycan is shown beyond this layer as a thin chain of alternating units of G l c N A c and M u r N A c. One unit of M u r N A c is connected to vertical chain of L A l a, D G l u, M A subscript 2 p m, D A l a from bottom to top. M A subscript 2 p m is single bonded to an N H subscript 3 group and single bonded to an N H group. The N H group is single bonded to Lysine, which is single bonded to C O O H and single bonded to murein lipoprotein, written parentheses aa close parentheses subscript 65. The murein lipoprotein is single bonded to Cysteine, which is single bonded to an N H group bound to the outer membrane and single bonded to a sulfur atom. The sulfur atom is single bonded to a C H subscript 2 group that is single bonded to C H that is single bonded to C H subscript 2. The C H is single bonded to an oxygen bound to the outer membrane and the terminal C H 2 is single bonded to an oxygen bound to the outer membrane. The inner edge of the outer membrane is a phospholipid layer, and the outer edge of the outer membrane is part of the L P S layer. The L P S layer consists of branching tendrils and M g superscript 3 plus.
A transmission electron micrograph of murein lipoprotein mutant. Two rod shaped bacteria are partially visible on the micrograph. The micrograph is centered on the point where the two bacteria meet. At this point, a circular line is visible between the two bacteria. The line is labeled, septation fails.
FIGURE 3.18 ■Gram-negative cell envelope (S-layer not shown).A. Murein lipoprotein has an N-terminal cysteine triglyceride inserted in the inward-facing leaflet of the outer membrane. The C-terminal lysine forms a peptide bond with the m-diaminopimelic acid (m-A2pm) of the peptidoglycan (murein) cell wall. Mg2⁺ ions cross-link the LPS chains. B. Lack of murein lipoprotein in mutant Salmonella causes the outer membrane to balloon out (arrow) when the cell tries to divide (TEM). JOAN FUNG ET AL. 1978. J. BACTERIOL.133:1467
Lipoprotein and lipopolysaccharide (LPS). In Gram-negative bacteria, the inward-facing leaflet of the outer membrane has a phospholipid composition similar to that of the inner membrane. The outer membrane’s inward-facing leaflet includes lipoproteins that connect the outer membrane to the peptide bridges of the cell wall. The major lipoprotein is called murein lipoprotein, also known as Braun lipoprotein (Fig. 3.18A). Murein lipoprotein consists of a protein with an N-terminal cysteine attached to three fatty acid side chains. The side chains are inserted in the inward-facing leaflet of the outer membrane. The C-terminal lysine forms a peptide bond with the m-diaminopimelic acid of peptidoglycan (murein). What happens to a mutant cell that fails to make murein lipoprotein? As the cell grows and divides, it fails to attach its outer membrane to the growing cell wall, causing the outer membrane to balloon out in the region where the daughter cells separate (Fig. 3.18B).
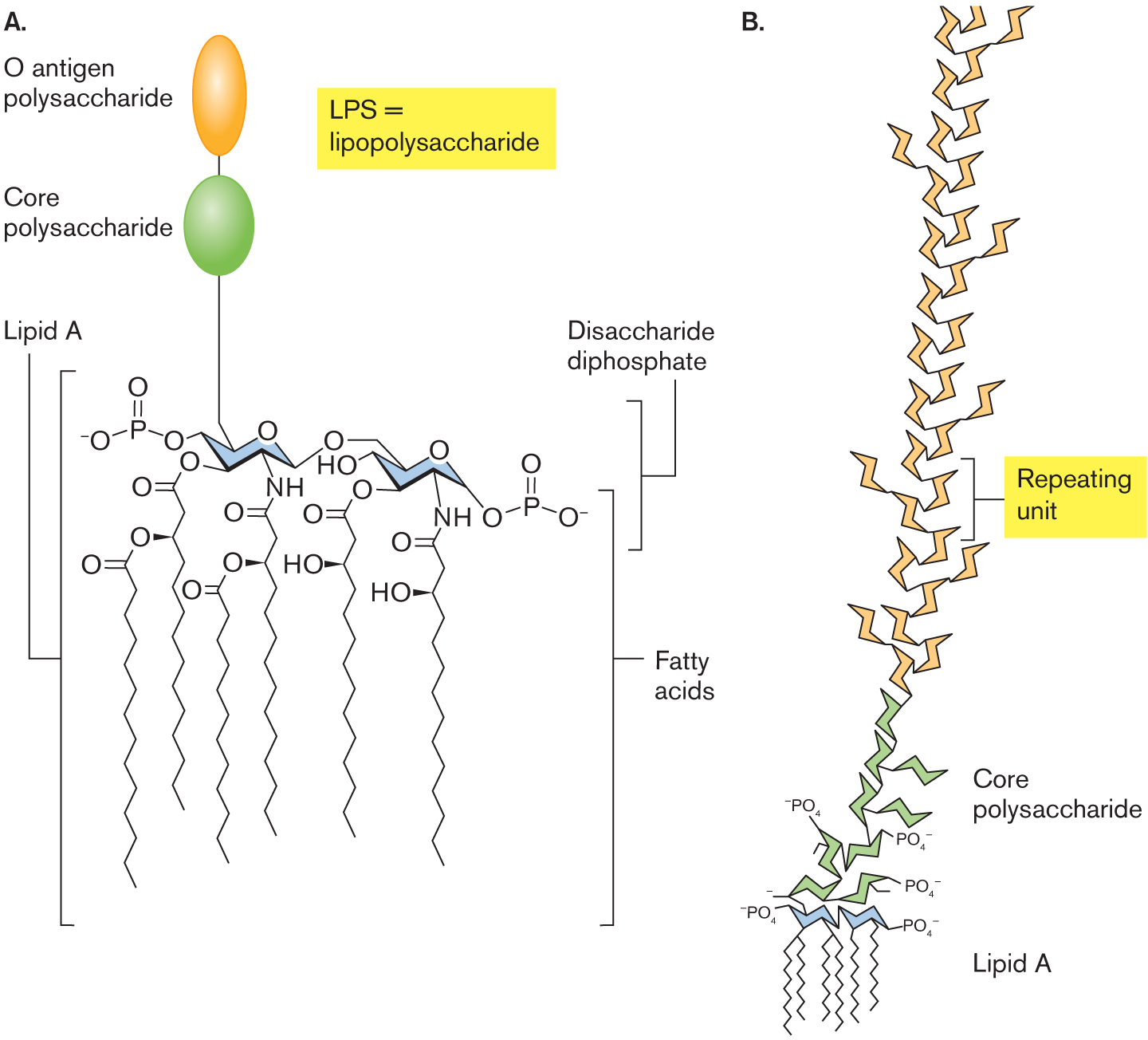
The outward-facing leaflet of the outer membrane has very different lipids from the inner leaflet. The main outward-facing phospholipids are called lipopolysaccharides (LPS; Fig. 3.19). LPS are of crucial medical importance because they act as endotoxins. Endotoxins are cell components that are harmless as long as the pathogen remains intact, but when released by a lysed cell, endotoxins overstimulate host defenses, inducing potentially lethal shock (discussed in Chapter 25). Thus, antibiotic treatment of an LPS-containing pathogen can kill the cells but can also lead to death of the patient.
More information
The molecular structure of lipopolysaccharide and an enlarged view of L P S.
The molecular structure of lipopolysaccharide Two chair configurations are shown. In the first configuration, C 1 is single bonded to oxygen and oxygen is single bonded to phosphate, in which phosphate is double-bonded to oxygen and single bonded to oxygen with minus ion, C 3 is substituted by oxygen, C 4 is single bonded to oxygen, C 5 is single bonded to N H, and C 6 is single bonded to oxygen. C 2 is bound to a core polysaccharide unit which is bound to an O antigen polysaccharide unit. In the second configuration, C 1 is single bonded to a hydroxyl group, C 2 is single bonded to oxygen, C 3 is substituted by oxygen, C 4 is single bonded to oxygen, in which oxygen is single bonded to phosphorous and phosphorous is double-bonded to oxygen at the top and single bonded to oxygen with minus ion, C 5 is single bonded to N H, and C 6 is single bonded to oxygen. These two models are attached to the six fatty acid tail at the bottom, which is double-bonded with oxygen and single bonded to the hydroxyl group. The chair configurations are placed within a bracket labeled Disaccharide diphosphate. The chair configurations bound to the fatty acid tails are placed within a bracket labeled Lipid A.
An illustration of the enlarged structure of L P S. Lipid A is bound to a core polysaccharide which is bound to repeating units of L P S.
FIGURE 3.19 ■Lipopolysaccharide (LPS).A. Lipopolysaccharide (LPS) consists of core polysaccharide and O antigen linked to a lipid A. Lipid A consists of a dimer of phosphoglucosamine esterified or amidated to six fatty acids. B. Repeating polysaccharide units of O antigen extend from lipid A.
The membrane-embedded anchor of LPS is lipid A, a molecule shaped like a six-legged giraffe (Fig. 3.19A). The lipid A moiety is the endotoxic part of LPS. The molecule’s six fatty acid “legs” have shorter chains than those of the fatty acids of the inner cell membrane, and two pairs are branched. The fatty acids have ester or amide links to the “body,” a dimer of glucosamine (an amino sugar also found in peptidoglycan). Analogous to the glycerol of glyceride phospholipids, each glucosamine has a phosphoryl group whose negative charge interacts with water. One glucosamine extends the long “neck” of the core polysaccharide, a sugar chain that reaches far outside the cell (Fig. 3.19B).
The core polysaccharide consists of five to ten sugars with side chains such as phosphoethanolamine. It extends to an O antigen, or O polysaccharide, a chain of as many as 200 sugars. The O polysaccharide may be longer than the cell itself. These chains of sugars form a layer that helps bacteria resist phagocytosis by white blood cells. The combination of sugar units in the O antigen varies greatly; E. coli clinical isolates show hundreds of different O-antigen sugar chains, which are recognized by distinct antibodies. Thus, O-antigen diversity offers one means by which pathogens evade the host immune system.
The LPS sugar side chains have negative charges that bind positively charged magnesium ions (Mg2+). Mg2+ ions thus form ionic cross-links that bridge LPS chains. The Mg2+-linked LPS can provide structural support for the cell, with strength comparable to that of the peptidoglycan layer.
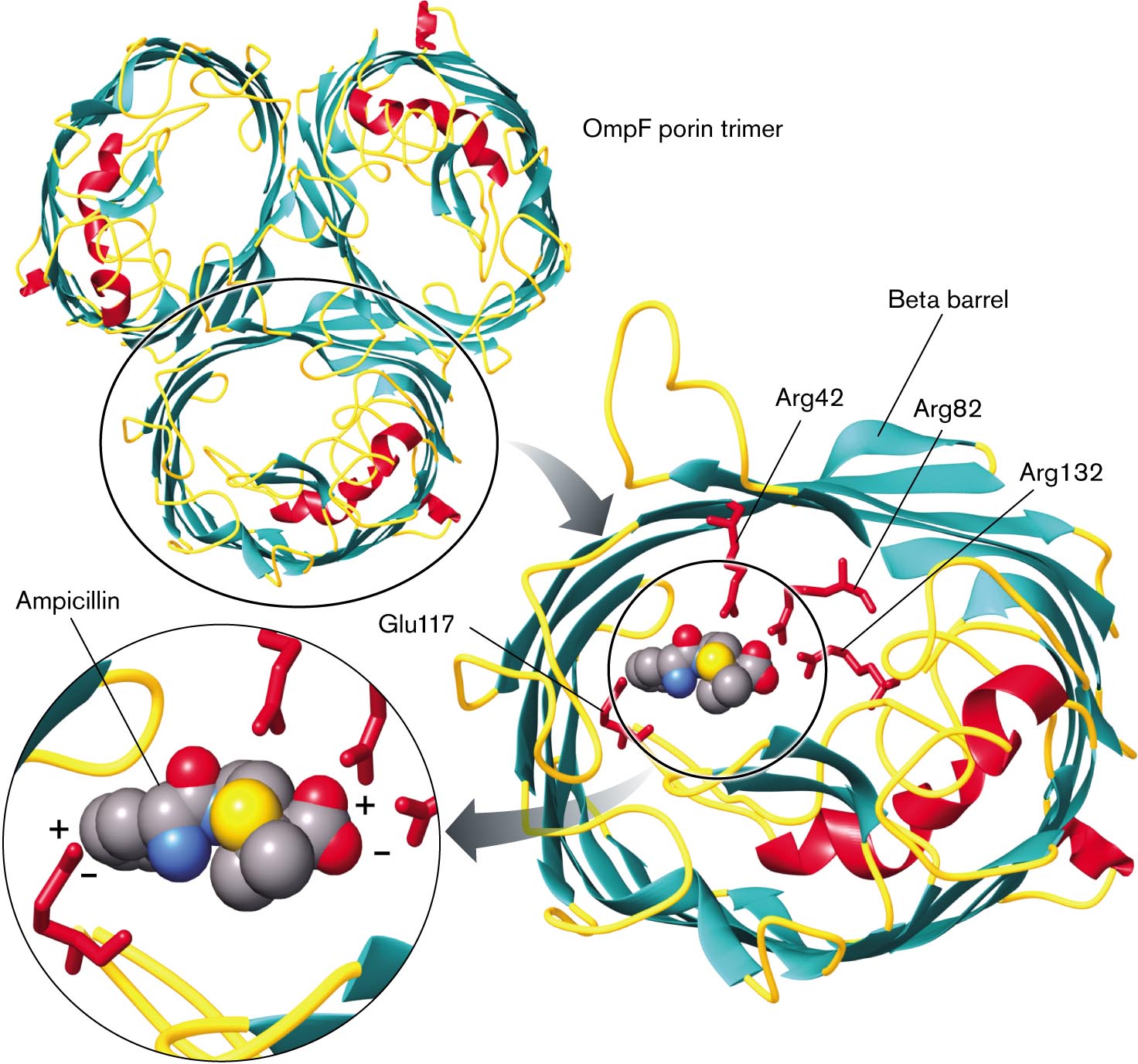
Outer membrane proteins. The outer membrane contains unique proteins not found in the inner membrane. Outer membranes contain a class of transporters called porins that permit the entry of nutrients such as sugars and peptides (Fig. 3.20). Outer membrane porins such as OmpF have a distinctive cylinder of beta sheet conformation (reviewed in eAppendix 1), also known as a beta barrel. A typical outer membrane porin exists as a trimer of beta barrels, each of which acts as a pore for nutrients.
More information
X-ray crystallography shows a model of transportation of ampicillin O m p f porin. It consists of three barrel-shaped beta-sheets bonded and labeled as O m p F porin trimer, in which one of the trimers is circled. An arrow leads to the magnified view of the O m p F bonded with four amino acids in the center, which is labeled as A r g 42, beta-barrel, A r g 82, Glu 117, and A r g 132. The inset view of the space-filling model several carbon atoms bonded with nitrogen, three oxygen, and nitrogen with plus and minus charge labeled as Ampicillin.
FIGURE 3.20 ■OmpF porin transports ampicillin. This model of the OmpF trimer is based on X-ray crystallography. (PDB code: 2OMF) Within each tubular porin monomer, charged amino acid residues contact ampicillin.
Source: Modified from Ekaterina M. Nestorovich et al. 2002. PNAS99:9789–9794.
Outer membrane porins have limited specificity, allowing passive uptake of various molecules—including antibiotics such as ampicillin. Ampicillin is a form of penicillin, which must get through the outer membrane to access the cell wall in order to block the formation of peptide cross-bridges. Ampicillin contains two charged groups and is thus unlikely to diffuse through a lipid bilayer. But the molecule crosses the E. coli outer membrane by passing through OmpF, where its charged groups are attracted to charged amino acid residues extending inside the porin (Fig. 3.20). Ampicillin’s positively charged amine group is attracted to the carboxylate of glutamate-117, and its negatively charged carboxylate is attracted to the arginine amines.
If porins can admit dangerous molecules as well as nutrients, should a cell make porins or not? In fact, cells express different outer membrane porins under different environmental conditions. In a dilute environment, cells express porins of large pore size, maximizing the uptake of nutrients. In a rich environment—for example, within a host—cells down-regulate the expression of large porins and express porins of smaller pore size, selecting only smaller nutrients and avoiding the uptake of toxins. For example, the porin regulation system of Gram-negative bacteria enables them to grow in the colon, which contains bile salts—a hostile environment for Gram-positive bacteria, which lack an outer membrane.
Periplasm. The outer membrane is porous to most ions and many small organic molecules, but it prevents the passage of proteins and other macromolecules. Thus, the region between the inner and outer membranes of Gram-negative cells, including the cell wall, defines a separate membrane-enclosed compartment of the cell known as the periplasm (see Fig. 3.18). The periplasm contains specific enzymes and nutrient transporters not found within the cytoplasm, such as periplasmic transporters for sugars, amino acids, or other nutrients. Periplasmic proteins are subjected to fluctuations in pH and salt concentration because the outer membrane is porous to ions. Some periplasmic proteins help refold proteins unfolded by oxidizing agents or by acidification.
Overall, the outer membrane, periplasm, inner membrane, and cytoplasm define four different cell compartments within a Gram-negative cell: two membrane-soluble compartments (outer and inner membranes), and two aqueous compartments (periplasm and cytoplasm). Each type of protein is typically found in only one of these locations. For example, the proton-translocating ATP synthase is found only in the inner membrane fractions, whereas sugar-accepting porins are only in the outer membrane.
Capsule. Some Gram-negative bacteria also form a capsule of loose glycolipids exterior to the outer membrane. For example, a capsule is found in virulent strains of Haemophilus influenzae, which was the leading cause of childhood meningitis before development of the Hib vaccine (see Chapter 24).
Thought Question
3.8 Why would proteins be confined to specific cell locations? Why would a protein not be able to function everywhere in the cell?
ANSWER ANSWER
Proteins have evolved one or more specific functions often optimized for a specific part of the cell. For example, water-conducting porins are found solely in the inner membrane (cell membrane), which is otherwise impermeable to water. The outer membrane, which is water permeable, is the sole location for specific porins that transport small peptides and sugars. The sugars then need to be taken across the inner membrane by transport proteins that have evolved to function best in this location. Similarly, different chaperones (proteins that aid peptide folding) have evolved to function best in the environment of the cytoplasm or periplasm, membrane-enclosed regions that differ substantially in pH and ion concentrations. In a different chemical environment of the cell, a protein may denature and lose its functional structure. A protein may be active only as part of a complex of proteins. If the protein is placed in a different location within the cell, its protein partners may be absent, rendering the protein nonfunctional.
Mycobacterial Cell Envelope
Exceptionally complex cell envelopes are found in Actinomycetes, a large and diverse phylum of soil bacteria that produce antibiotics and other industrially useful products (discussed in Chapter 18). The most complex envelopes known are those of actinomycete-related bacteria, the mycobacteria. Mycobacteria include the famous pathogens Mycobacterium tuberculosis (the cause of tuberculosis) and M. leprae (the cause of leprosy). Their complex envelope prevents use of the Gram stain. The mycobacterial envelope may be 1,000-fold less permeable to nutrients and toxins than the envelope of E. coli is. Thus, mycobacteria must grow slowly—but they effectively resist host defenses.
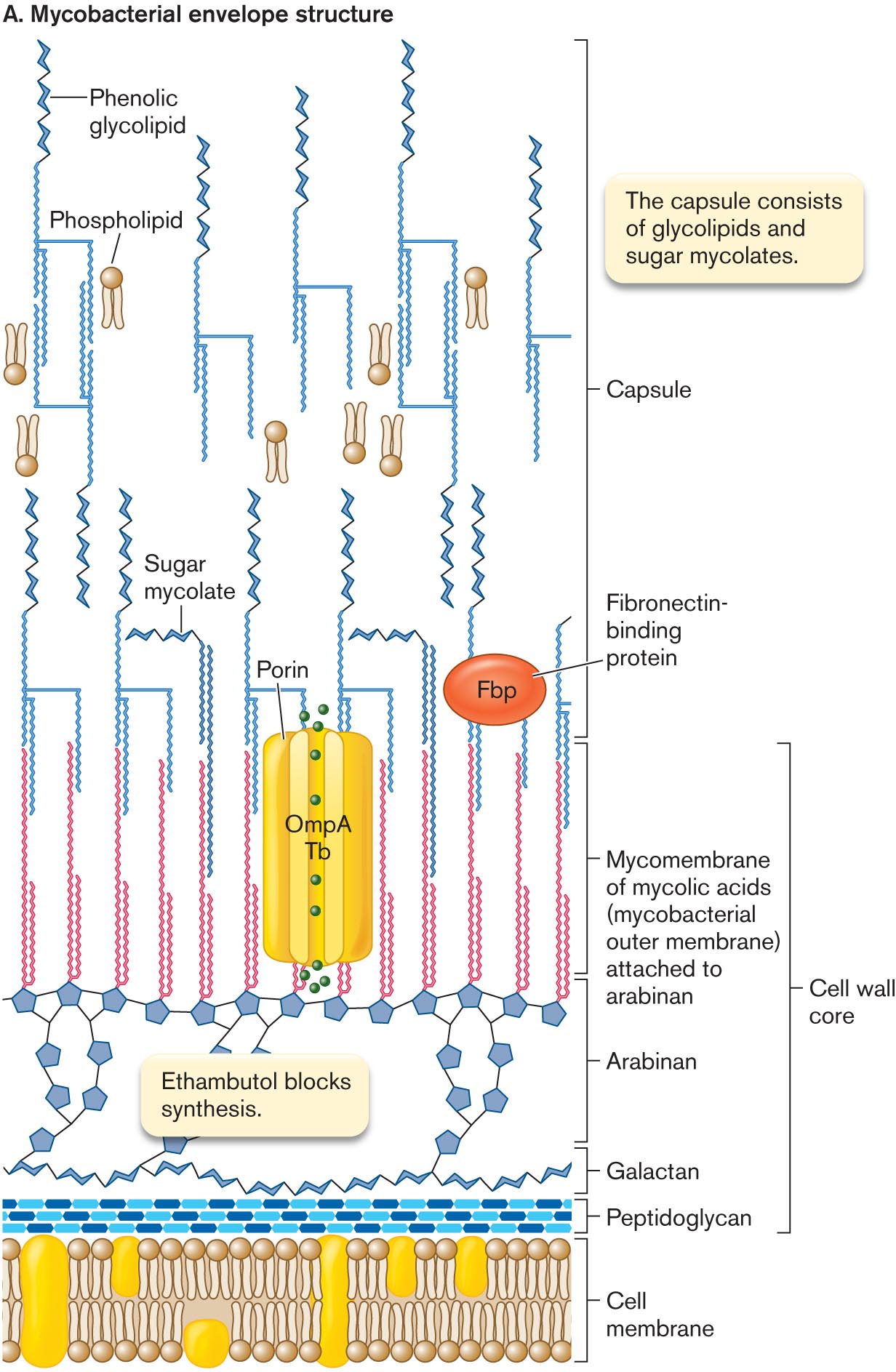
The mycobacterial envelope includes features of both Gram-positive and Gram-negative cells, as well as structures unique to mycobacteria (Fig. 3.21). In mycobacteria, the peptidoglycan is linked to chains of galactose, called galactans. The galactans are attached to arabinans, polymers of the five-carbon sugar arabinose. The arabinan-galactan polymers are known as arabinogalactans. Arabinogalactan biosynthesis is inhibited by two major classes of anti-tuberculosis drugs: ethambutol and the benzothiazinones.
More information
An illustration of mycobacterial envelope structure and a cryo- electron micrograph of a cell membrane.
An illustration of mycobacterial envelope structure. Several cyclohexanes bonded at the top of the cell membrane in three rows are labeled as peptidoglycan. The several chair conformation model bonded in the sequence above the peptidoglycan is labeled as galactan, which is connected to several five-membered carbon rings labeled as Ethambutol blocks synthesis is labeled as Arabian. The double-stranded vertical chains are bonded at the top of each five-membered carbon ring, and the corresponding text reads Mycomembrane of mycolic acids, or mycobacterial outer Membrane, attached to arabinan, which is labeled as cell wall core. It is bonded with the structure of porin in the center is labeled as O m p A T b and an oval-shaped image labeled as Fibronectin- binding protein. The sugar mycolate is attached to the top of the mycomembrane acids, which is further connected to the Phenolic glycolipid at the top, and the symbol of phospholipid is placed beside, is labeled as a capsule. The corresponding text reads the capsule consists of glycolipids and sugar mycolates.
More information
A cryo- transmission electron micrograph of a mycobacterial cell envelope. A half circle of a cell is visible. In this view, the cell has a radius of about 100 nanometers. The cell membrane is identified. A thick layer exterior the cell membrane is identified as the mycomembrane.
FIGURE 3.21 ■Mycobacterial envelope structure.A. The peptidoglycan layer is linked to a chain of galactose polymer (galactan) and arabinose polymer (arabinan). Arabinan forms ester links to mycolic acids, which form an outer bilayer with phenolic glycolipids. B. Mycobacterial envelope (cryo-TEM). B. CHRISTIAN HOFFMAN ET AL. 2008. PNAS105:3963
The ends of the arabinan chains form ester links to mycolic acids (uncharged mycolates). Mycolic acids provide the basis for acid-fast staining, in which cells retain the dye carbolfuchsin, an important diagnostic test for mycobacteria and actinomycetes (described in Chapter 28). Mycolic acids contain a hydroxy acid backbone with two hydrocarbon chains—one comparable in length to typical membrane lipids (about 20 carbons), the other about threefold longer. The long chain includes ketones, methoxyl groups, and cyclopropane rings. Hundreds of different forms are known.
The mycolic acids form a bilayer interleaved with sugar mycolates—a kind of outer membrane, or “mycomembrane,” analogous to the Gram-negative outer membrane. This mycomembrane even contains porins homologous to Gram-negative beta barrel porins such as OmpA. Other mycolate-embedded proteins include virulence factors such as fibronectin-binding protein (Fbp). Fbp enhances the ability of M. tuberculosis to invade macrophages.
The outer ends of the sugar mycolates are interleaved with phenolic glycolipids, which include phenol groups linked to sugar chains. The extreme hydrophobicity of the phenol derivatives generates a waxy surface that prevents phagocytosis by macrophages. Overall, the thick, waxy envelope excludes many antibiotics and offers exceptional protection from host defenses, enabling the pathogens of tuberculosis and leprosy to colonize their hosts over long periods. However, the thick envelope also retards uptake of nutrients. As a result, M. tuberculosis and M. leprae grow extremely slowly and are a challenge to culture in the laboratory.
Bacterial Cytoskeleton
In eukaryotes, cell shape has long been known to be maintained by a cytoskeleton of protein microtubules and filaments (reviewed in eAppendix 2). But what determines the shape of bacteria? We saw earlier that bacterial shape is in part maintained by the cell wall and the resulting turgor pressure. But research over the past decade shows that bacteria also possess protein cytoskeletal components—and remarkably, some of them are homologous to eukaryotic cytoskeletal proteins. For example, the MreB tracker protein for peptidoglycan synthesis (Fig. 3.13) is a part of the bacterial cytoskeleton. MreB is a homolog of the eukaryotic microfilament protein actin.
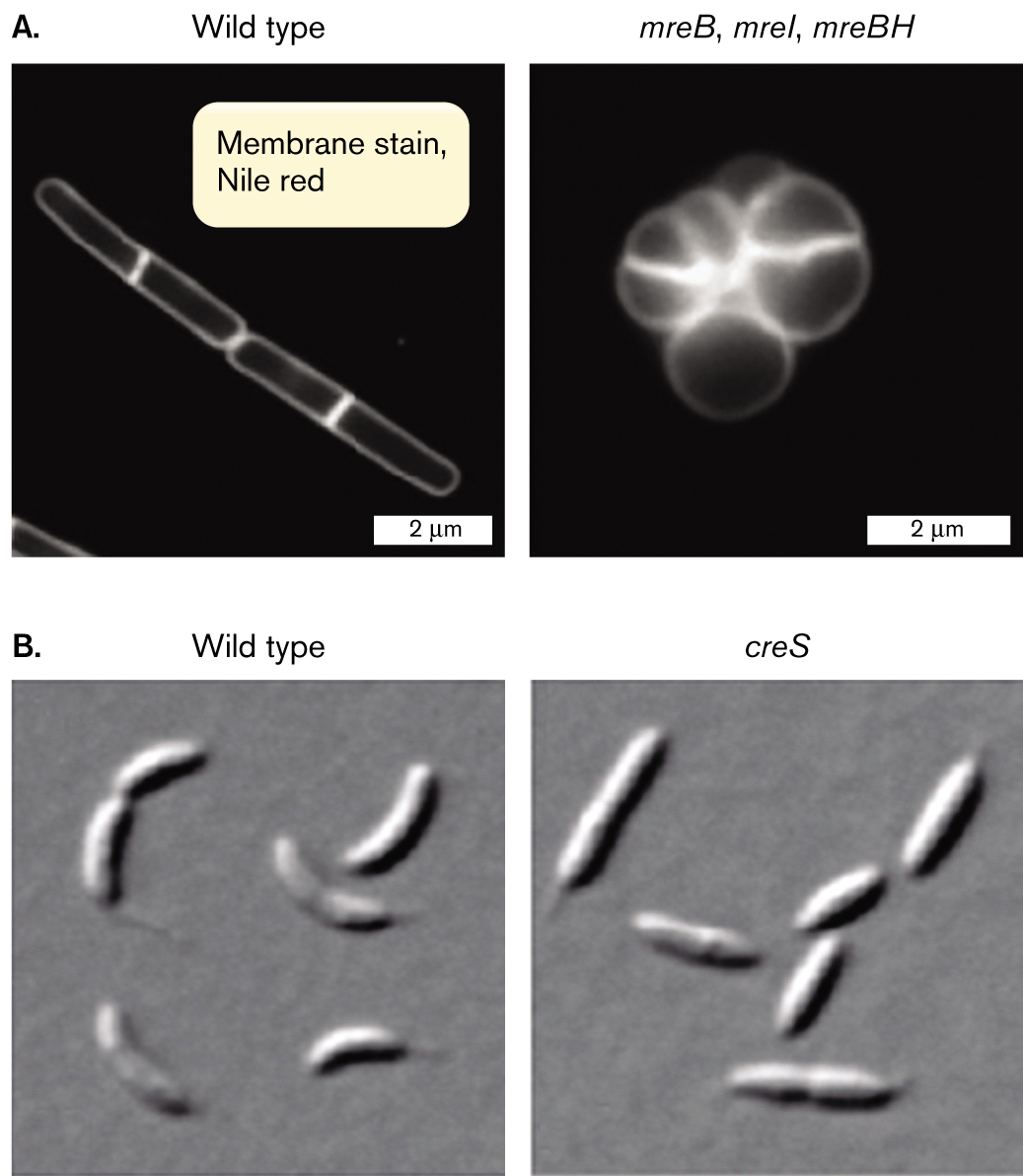
The bacterial cytoskeletal proteins are revealed by gene defects that drastically alter the cell shape. For example, Figure 3.22A compares wild-type cells of Bacillus subtilis with cells containing mutations in three mreB homologs (mreB, mreI, and mreBH). The wild-type cells have a defined rod shape, whereas the mutant shape is round and undefined. The mutant lacks the MreB complex that regulates peptidoglycan synthesis and thereby defines the rod-shaped cell. Another example of a shape-altering mutation affects the comma-shaped cell of Caulobacter crescentus (Fig. 3.22B). A mutation in the gene creS results in cells that are straight instead of curved. The creS gene expresses the cytoskeletal protein CreS (crescentin).
More information
Four electron micrographs comparing wild type cells to cytoskeletal mutant cells.
An electron micrograph of wild type Bacillus subtilis compared to a cytoskeletal mutant. The wild type micrograph shows two rod shaped cells of 4 micrometers in length and 0.5 micrometer in width. A bright band is visible in the center of each cell. Overlaying text reads, membrane stain Nile red. The mutant micrograph is labeled m r e B, m r e I, m r e B H. A cluster of faint circular shapes is visible. Bright bands run erratically through the cluster. The cluster makes a rough circle of about 2 micrometers in diameter.
An electron micrograph of wild type Caulobacter crescentus compared to a cytoskeletal mutant. The wild type micrograph shows several rod shaped bacteria, with slight bends in their centers. The mutant micrograph is labeled c r e S. The micrograph shows several rod shaped bacteria with no bend.
FIGURE 3.22 ■Cytoskeletal mutants. Wild-type cells compared with mutants in (A)Bacillus subtilis (fluorescence microscopy) and (B)Caulobacter crescentus (DIC).
Source: Part A from https://www.nature.com/articles/ncomms4442; https://creative commons.org/license/by/3.0/us/legalcode.
T. NILSEN ET AL. 2005. J. BACTERIOL.187:6187M. CABEEN ET AL. 2010. J. BACTERIOL.192:3368
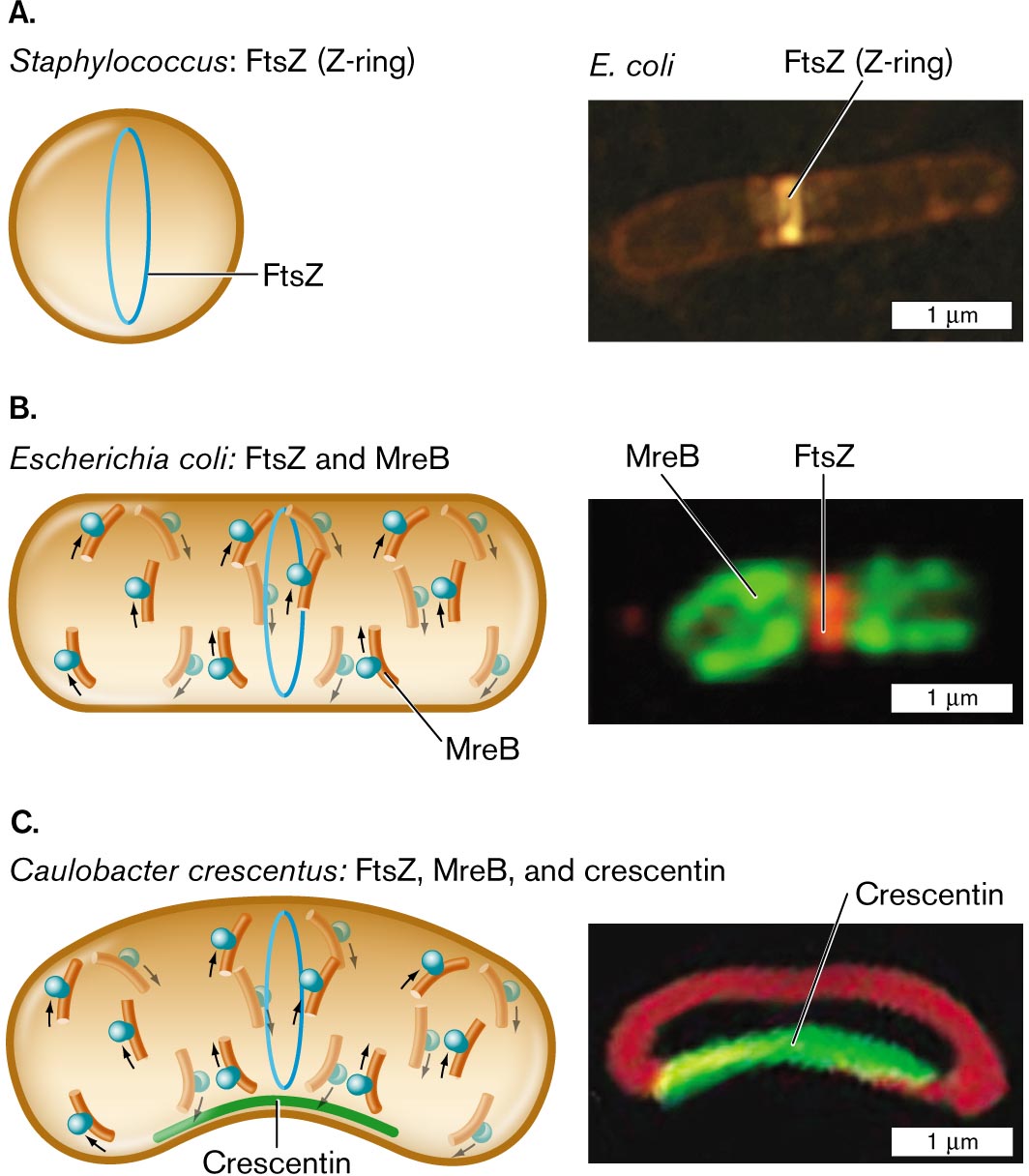
How do the various cytoskeletal proteins work together to generate the overall shape of a bacterial cell? The functions of cytoskeletal proteins are probed by fluorescent protein fusions (Fig. 3.23). In both spherical bacteria (cocci) and rod-shaped bacilli, cell division requires the protein FtsZ, a homolog of the eukaryotic protein tubulin (the subunit of eukaryotic microtubules). The bacterial protein FtsZ forms a ring-shaped complex around the middle of the cell, called the Z-ring. The Z-ring determines the cell diameter and manages the growth of the dividing partition, which is called the septum (plural, septa).
For a rod-shaped cell, elongation requires polymerization of MreB (Fig. 3.23B). MreB travels in a helical arc beneath the cell membrane, guiding peptidoglycan elongation. If the rod shape is curved (forming a crescent shape), the third cytoskeletal protein, crescentin, polymerizes along the inner curve of the crescent (Fig. 3.23C). The cell’s outer curve is visualized by a membrane-specific fluorophore. These cytoskeletal proteins, and their variants that have evolved in other species, work together within cells to generate the shapes of bacteria.
More information
Three illustrations and accompanying fluorescence micrographs describing shape determining proteins in different bacteria.
An illustration and an accompanying fluorescence micrograph of F t s Z in Staphylococcus. The cell is circular shaped, and inside the cell is a vertical oval ring labeled F t s Z. To the right is a microscopic image of F t s Z, or Z-ring, fluorescing in an E. coli bacterium. E. coli is a rod shaped cell of about 3 micrometers in length and 0.5 micrometer in width. F t s Z is identified as a bright band near the center of the cell.
An illustration and an accompanying fluorescence micrograph of F t s Z and M r e B in Escherichia coli. The cell is a long horizontal rod shape. The F t s Z ring is in the center, and overlapping and to the left and right are M r e B rings. To the right shows a microscopic fluorescent image of F t s Z bonded in the center of a cell and M r e B at the edges. The unit is about 2 micrometers long and 1 micrometer wide.
An illustration and an accompanying fluorescence micrograph of F t s Z, M r e B, and crescentin in Caulobacter crescentus. The cell is a long horizontal rod shape with a central bend. The bottom part of the cell where it curves is labeled Crescentin. F t s Z ring is located in the center of the cell and overlapping, and to the left and right are M r e B rings. To the right is a microscopic fluorescent image of a rod shaped cell with a central bend. The bend is fluorescent and labeled Crescentin. The cell is about 2 micrometers long and 1 micrometer wide.
FIGURE 3.23 ■Shape-determining proteins.A. Cell diameter is maintained by FtsZ polymerization to form the Z-ring. B. Elongation of a rod-shaped cell requires MreB proteins. MreB polymerizes around an E. coli cell (MreB-YFP fluorescence) along with a Z-ring of FtsZ (fluorescent anti-FtsZ antibody). C. Crescent-shaped cells possess a third shape-determining protein, CreS (crescentin), which polymerizes along the inner curve of the crescent. Crescentin protein fused to green fluorescent protein (CreS-GFP) localizes to the inner curve of Caulobacter crescentus. Membrane-specific stain FM4-64 (red fluorescence) localizes to the membrane around the cell. Q. SUN AND W. MARGOLIN. 1998. J. BACTERIOL.180:2050PURVA VATS AND LAWRENCE ROTHFIELD. 2007. PNAS104:17795NORA AUSMEES ET AL. 2003. CELL115:705
To Summarize
The cell wall maintains turgor pressure. The cell wall is porous, but its network of covalent bonds generates turgor pressure that protects the cell from osmotic shock.
The Gram-positive cell envelope has multiple layers of peptidoglycan, threaded by teichoic acids.
The protein S-layer of Gram-negative and Gram-positive bacteria is highly porous but can prevent phagocytosis and protect cells in extreme environments. In many archaea, the S-layer serves the structural function of a cell wall.
The capsule, composed of polysaccharide and glycoprotein filaments, protects cells from phagocytosis. Both Gram-positive and Gram-negative cells may possess a capsule.
The Gram-negative outer membrane regulates nutrient uptake and excludes toxins. The outer membrane contains LPS and protein porins of varying selectivity.
The mycobacterial cell wall includes features of both Gram-positive and Gram-negative cells. The arabinogalactan layer adds thickness to the cell wall. The mycolate outer membrane and phenolic glycolipids limit uptake of nutrients and antibiotics.
The bacterial cytoskeleton includes proteins that regulate cell size, play a role in determining the rod shape of bacilli, and generate curvature in crescent-shaped cells.
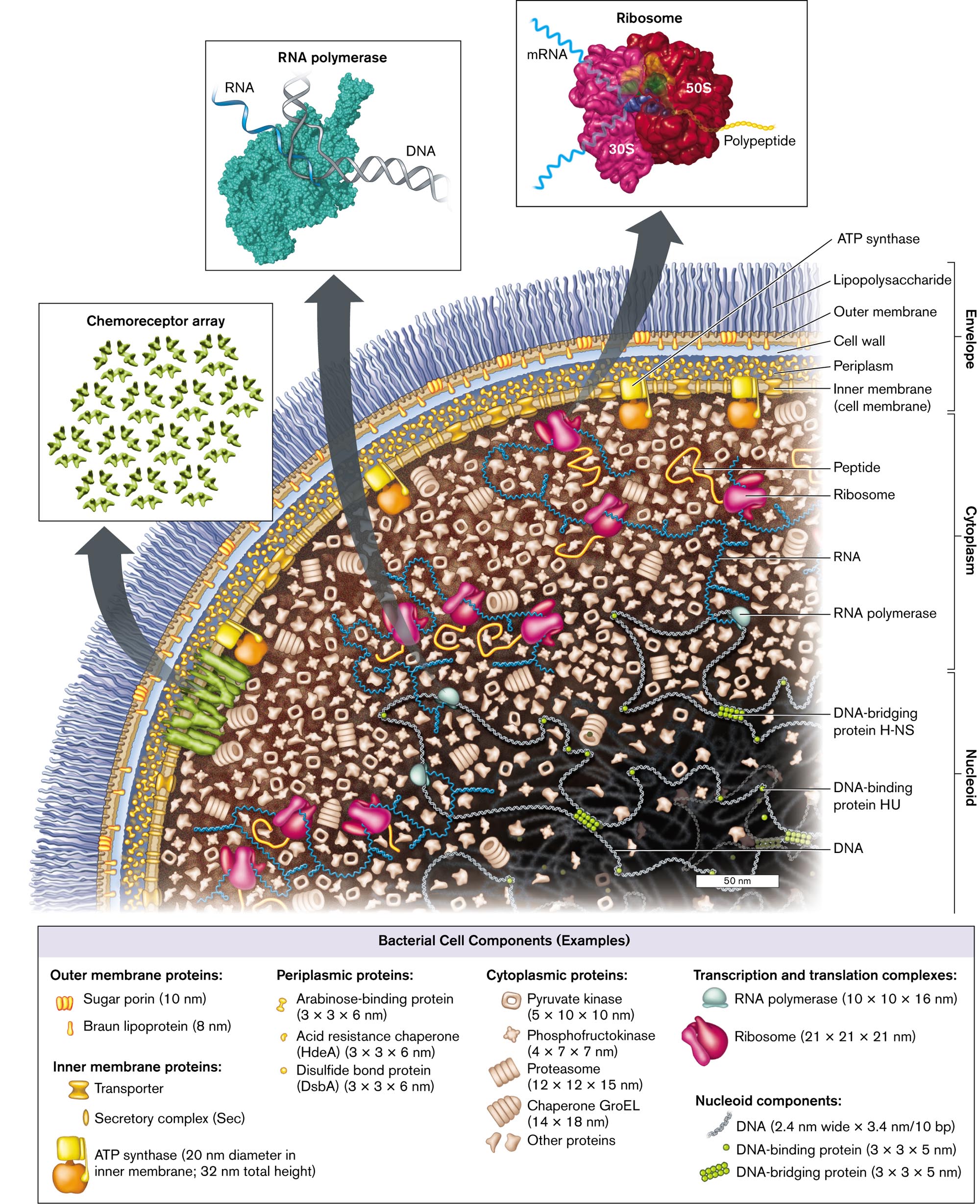
An illustration of the cell components and structure of Escherichia coli. The bacterium consists of small molecules such as nucleoid, cytoplasm, and envelope. The nucleoid contains D N A, D N A-binding protein H U, and D N A-bridging protein H-N S. The cytoplasm contains R N A polymerase, R N A, peptides, and ribosomes. The envelope contains the inner Membrane, or cell membrane, periplasm, the cell wall, the Outer Membrane, and lipopolysaccharides. A flagellar motor connects to the inner Membrane and travels outward. An inset shows a model of an R N A polymerase with the labeled parts D N A and R N A. Another inset from the bacterium shows a model of several D N A-binding proteins is labeled as Chemoreceptor array. The third inset shows a model of Ribosome bonded with m R N A, 30 S, 50 S, and polypeptide chain at a magnification of 50 nanometers. The fourth inset shows a microscopic view of the whole bacterium, and the labeled parts are the envelope, the nucleoid, and ribosomes at a magnification of 0.25 micrometers. A box containing examples of bacterial cell components is as follows: Outer membrane proteins: Sugar porin, 10 nanometers long, and Braun lipoprotein, 8 nanometers long; Inner membrane proteins: A T P synthase, 20 nanometers diameter in the inner Membrane; 32 nanometers total height, Secretory complex, abbreviated S e c, and Transporter. Periplasmic proteins: Arabinose-binding protein, dimensions 3 times 3 times 6 nanometers, Disulfide bond protein, abbreviated D s b A, dimensions 3 times 3 times 6 nanometers, Acid resistance chaperone, abbreviated H d e A, dimensions 3 times 3 times 6 nanometers. Cytoplasmic proteins: Pyruvate kinase, dimensions 5 times 10 times 10 nanometers, Phosphofructokinase, dimensions 4 times 7 times 7 nanometers, Proteasome, dimensions 12 times 12 times 15 nanometers, Chaperone G r o E L, dimensions 14 times 18 nanometers, Other proteins. Transcription and translation complexes: R N A polymerase, dimensions 10 times 10 times 16 nanometers, Ribosome dimensions 21 times 21 times 21 nanometers. Nucleoid components: D N A, dimensions 2.4 nanometers wide times 3.4 nanometers per 10 b p, D N A-binding protein, dimensions 3 times 3 times 5 nanometers, and D N A-bridging protein, dimensions 3 times 3 times 5 nanometers.
FIGURE 3.1 ■Escherichia coli: a Gram-negative bacterium of the gut microbiome. The envelope includes the outer membrane, the cell wall and periplasm, and the inner (cell) membrane with an embedded chemoreceptor array. The cytoplasmcontains enzymes, messenger RNA (mRNA) extending out of the nucleoid, and ribosomes. Ribosomes translate the mRNA to make proteins, which are folded by chaperones. The nucleoidcontains the chromosomal DNA wrapped around binding proteins. (PDB codes: ribosome, 1GIX, 1GIY; RNA polymerase, 1MSW)
An illustration of the Gram-negative cell envelope structure and a micrograph of a murein lipoprotein mutant.
An illustration of the Gram-negative cell envelope structure. The inner cell membrane is shown as a phospholipid bilayer. Peptidoglycan is shown beyond this layer as a thin chain of alternating units of G l c N A c and M u r N A c. One unit of M u r N A c is connected to vertical chain of L A l a, D G l u, M A subscript 2 p m, D A l a from bottom to top. M A subscript 2 p m is single bonded to an N H subscript 3 group and single bonded to an N H group. The N H group is single bonded to Lysine, which is single bonded to C O O H and single bonded to murein lipoprotein, written parentheses aa close parentheses subscript 65. The murein lipoprotein is single bonded to Cysteine, which is single bonded to an N H group bound to the outer membrane and single bonded to a sulfur atom. The sulfur atom is single bonded to a C H subscript 2 group that is single bonded to C H that is single bonded to C H subscript 2. The C H is single bonded to an oxygen bound to the outer membrane and the terminal C H 2 is single bonded to an oxygen bound to the outer membrane. The inner edge of the outer membrane is a phospholipid layer, and the outer edge of the outer membrane is part of the L P S layer. The L P S layer consists of branching tendrils and M g superscript 3 plus.
A transmission electron micrograph of murein lipoprotein mutant. Two rod shaped bacteria are partially visible on the micrograph. The micrograph is centered on the point where the two bacteria meet. At this point, a circular line is visible between the two bacteria. The line is labeled, septation fails.
FIGURE 3.18 ■Gram-negative cell envelope (S-layer not shown).A. Murein lipoprotein has an N-terminal cysteine triglyceride inserted in the inward-facing leaflet of the outer membrane. The C-terminal lysine forms a peptide bond with the m-diaminopimelic acid (m-A2pm) of the peptidoglycan (murein) cell wall. Mg2⁺ ions cross-link the LPS chains. B. Lack of murein lipoprotein in mutant Salmonella causes the outer membrane to balloon out (arrow) when the cell tries to divide (TEM). JOAN FUNG ET AL. 1978. J. BACTERIOL.133:1467
A detailed illustration of peptidoglycan synthesis. Synthesis is shown to take place between the outer membrane and the inner membrane. Synthesis is mediated by a peptidoglycan synthesis complex, shown blown-up in the illustration. The complex extends from the interior of the cell across the inner membrane into the space between the inner and outer membranes. An arch shaped structure supports the complex, the structure is comprised of M r e B patches. Attached to the arch are several irregularly shaped proteins labeled M r e D, M r e C, R o d A, and R o d Z. Two penicillin binding proteins sit atop M r e C. The penicillin binding proteins are labeled P B P 2 and P B P 1 A. A chain of peptidoglycan is synthesized through the complex.
FIGURE 3.13 ■Peptidoglycan synthesis is organized by penicillin-binding proteins (PBP2, PBP1A) and by cytoskeletal proteins. Protein MreB guides the direction of synthesis in helical arcs around the cell.
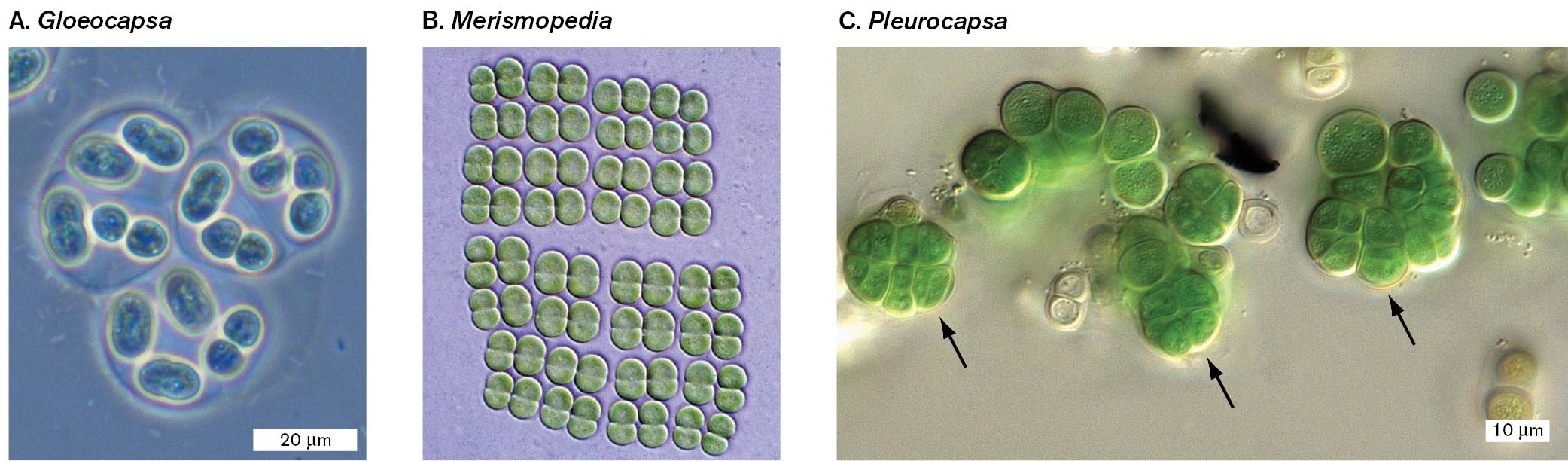
Three micrographs of the structures of colonial cyanobacteria. Part A is a micrograph of Gloeocapsa cells surrounded by mucus. Several oval shaped cells can be seen grouped into sections. Each section is surrounded by a thin layer of mucus. Part B is a micrograph of Merismopedia cells arranged into extended quartets. Many dicocci cells are visible. The cells are arranged into quartets and further arranged into octets. Part C is a micrograph of Pleurocapsa cells with baocytes formations. There are many cocci cells in the micrograph. Some of the cells are stacked so that they have a clover like appearance. These stacked cells are the baocytes formations.
A micrograph of Gloeocapsa cells surrounded by mucus. Several small oval cells are visible. Each cell is about 5 micrometers wide and between 5 to 10 micrometers long. The cells are grouped into 3 sections. Each section is surrounded by a thin layer of mucus. The mucus sections are roughly circular in shape and are about 20 micrometers in diameter.
A micrograph of Merismopedia cells arranged into extended quartets. Many dicocci Merismopedia cells can be seen. The cells appear green in the micrograph. The cells are arranged into quartets that are further arranged into octets. There is no scale provided.
A micrograph of Pleurocapsa cells with baocytes formations. Many cocci cells are visible. They appear green in the micrograph. The cells are each about 10 micrometers in diameter. Some of the cells are stacked on top of each other which gives a four-leaf clover like appearance. Arrows indicate that these stacked cells are baocytes formations.
FIGURE 18.8 ■Colonial cyanobacteria.A.Gloeocapsa is surrounded by mucus. Cells grow as single cells, doublets, or quartets. B.Merismopedia forms extended quartets, octets, and so on. C. Unicellular cyanobacterium Chroococcidiopsis sp. with baocytes (arrows) formed by successive multiple divisions. MICHAEL ABBEY/SCIENCE SOURCEMICHAEL ABBEY/SCIENCE SOURCEBURKHARD BÜDEL
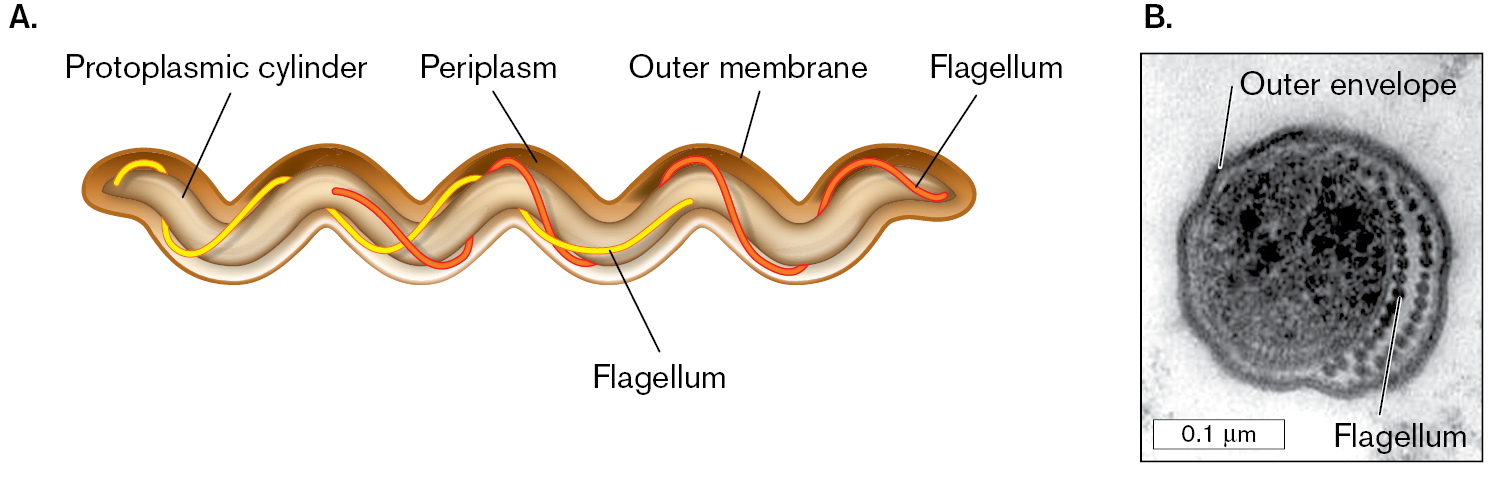
A diagram and a micrograph of spirochete cell structure. Part A is the diagram of spirochete cell structure. The protoplasmic cylinder is the main wavy, tubular structure of the cell. The outer membrane surrounds the protoplasmic cylinder, tracing the wave shape. Periplasm is between the protoplasmic cylinder and the outer membrane. Flagella wrap around the protoplasmic cylinder in a spiral. Part B is a transmission electron micrograph of a cross-section of a human gingival spirochete. The cross section is round. The outer envelope is a thick dark ring around the cell. To the right of the circle within the outer envelope is a chain of dots labeled flagellum. The protoplasmic cylinder is a dark, grainy circle of about 0.1 micrometer in diameter to the left of the flagellum.
A diagram of spirochete cell structure. The protoplasmic cylinder is the main wavy, tubular structure of the cell. The outer membrane surrounds the protoplasmic cylinder, tracing the wave shape. Periplasm is between the protoplasmic cylinder and the outer membrane. Flagella wrap around the protoplasmic cylinder in a spiral.
A transmission electron micrograph of a cross-section of a human gingival spirochete. The micrograph is in greyscale. The cross section is round. It has a diameter of 0.2 micrometer. The outer envelope is a thick dark ring around the cell. To the right of the circle within the outer envelope is a chain of dots labeled flagellum. The protoplasmic cylinder is a dark, grainy circle of about 0.1 micrometer in diameter to the left of the flagellum.
FIGURE 18.43 ■Spirochete structure.A. Spirochete cell structure, showing the arrangement of periplasmic flagella. B. Cross section through a human gingival spirochete, showing outer envelope and flagella (axial fibrils; TEM). M. A. LISTGARTEN AND S. S. SOCRANSKY. 1964. J. BACTERIOL.88:1087
 ANSWER
ANSWER ANSWER
ANSWER