3.5 Cell Asymmetry, Membrane Vesicles, and Extensions
Do dividing bacteria produce symmetrical offspring? Even superficially symmetrical bacilli such as Escherichia coli show underlying chemical and physical asymmetry, such as possession of a chemoreceptor array at the “forward” pole. Other species, such as Caulobacter crescentus, develop different structures at either pole, and their cell division generates two different cell types. And many kinds of bacteria extend their cytoplasm in surprising ways, by forming extracellular membrane vesicles and extensions that may interact with other cells. Such cell extensions complicate the very definition of an individual cell.
Bacterial Cell Differentiation
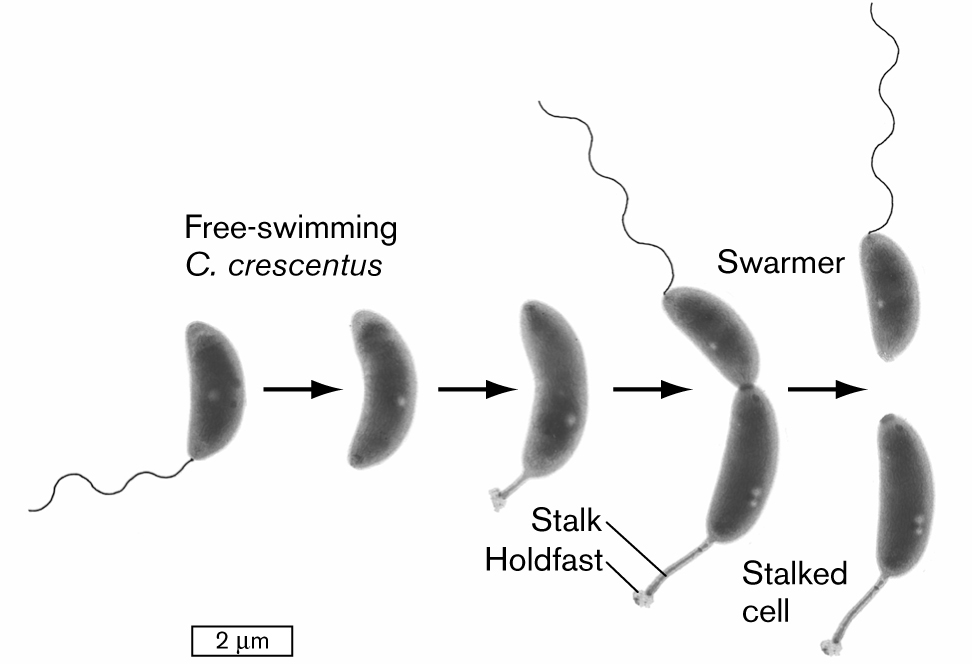
Bacteria whose poles have different structures generate two different forms of progeny. The wetland bacterium Caulobacter crescentus has one plain pole and one pole with either a flagellum or a cytoplasmic extension called a stalk (Fig. 3.30). A flagellated cell (also known as a swarmer cell) swims freely in an aqueous habitat, such as a pond or a sewage bed. After swimming for about half an hour, if the bacterium finds a place with enough nutrients, the cell sheds its flagellum and replaces it with a stalk. The stalked cell attaches to sediment and then immediately starts to replicate its DNA and divides, producing a flagellated daughter cell, as well as a daughter cell containing the original stalk.
More information
An illustration shows a developmental model for the division of an asymmetrical cell. It begins with a swarmer cell, which is a bean-shaped structure attached with a tail that is labeled as free-swimming C. crescentus. It leads to the next two images. The cell continues to elongate. There is a flagellum at one end of the cell and a stalk with a holdfast on the other end of the cell. The cell is pinched in the middle but still connected and leads to the division of two cells, in which one cell is attached with tail is labeled as a swarmer, and another cell is labeled as the stalked cell. The cell is about 2 micrometers long at the start and elongates to about 3 micrometers in length.
FIGURE 3.30 ■Asymmetrical cell division: a model for development. A swarmer cell of Caulobacter crescentus loses its flagellum and grows a stalk. The stalked cell divides to produce a swarmer cell (TEM). YVES BRUN
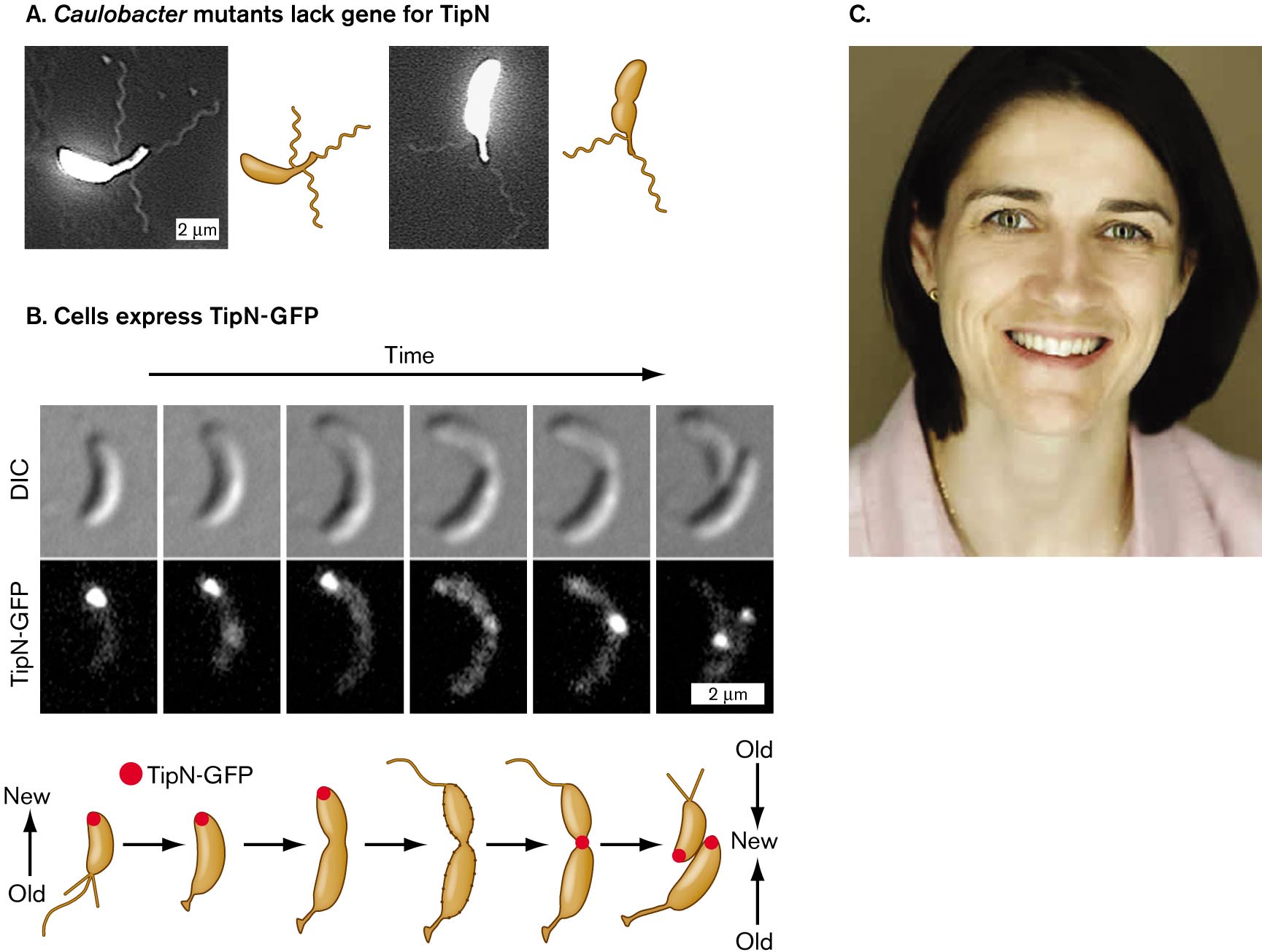
How does C. crescentus organize itself to produce two different cell types, each with a different organelle at one pole? The process is a rudimentary form of cell differentiation, comparable to the differentiation processes that animal cells undergo in the embryo. The C. crescentus life cycle is governed by regulator proteins such as TipN, studied by students of Christine Jacobs-Wagner at Yale University. Mutants lacking TipN make serious mistakes in development. Instead of making a single flagellum at the correct cell pole, the cell makes multiple flagella at various locations, even on the stalk (Fig. 3.31A). Jacobs-Wagner proposed that TipN is a landmark protein that correctly marks the site of a new cell pole and directs the polar placement of flagella.
Figure 3.31B shows cells expressing TipN fused to the fluorescent protein GFP, which is then detected by fluorescence microscopy. (A gene fusion expresses fused proteins that fluoresce but may still perform the original protein’s function, as shown in Figure 2.29.) The fluorescent fusion protein TipN-GFP localizes to the cell pole opposite the stalk. As the cell prepares to divide, TipN leaves the pole, delocalizing around the cell. Eventually, the TipN protein relocalizes at the septum, where the new poles appear.
More information
Micrographs showing the role of T i p N protein in Caulobacter, and a photo of Christine Jacobs-Wagner.
Micrographs of Caulobacter mutants that lack the gene for T i p N. In the first micrograph, a bean shaped bacterium has three flagella. The bacterium is about 4 micrometers long. In the second micrograph, a bean shaped bacterium has two flagella. The bacterium is about 4 micrometers long.
An illustration shows cells that express T i p N-G F P. There are three rows of a cell dividing over time. The first row is called D I C, and there are six images of a cell dividing into two. There is an arrow pointing at the bottom of the cell in each image. The second row is called T i p N-G F P, and there are six images of a cell dividing into two. There is a large glowing mass that moves in each image. In the first image, the mass is at the top of the cell. In the second image, the mass is at the top of the elongated cell. In the third image, the mass is at the very top of the elongated cell. In the fourth image, the mass is not visible. In the fifth image, the mass is in the center of the cell. In the sixth image, the mass has split into two, and the light is at the top of the two newly split cells are shown at a magnification of 2 micrometers. Below is an illustration of the six steps of the cell dividing into two cells. It begins from a cell with flagella and a mass of T i p N-G F P at the top of the cell. The bottom of the cell is old, and the top of the cell is new. In the next step, the cell has no flagella but is starting to grow a stalk, and the T i p N-G F P is still at the top. In the third step, the cell is starting to pinch in the center. In the fourth step, there is no mass of T i p N-G F P, and there is a flagellum at the top of the cell and a stalk and holdfast on the bottom of the cell. In the fifth step, the T i p N-G F P is in the center of the cell. In the last image, there are two cells with a mass of T i p N-G F P at the top of each new cell. The cells are facing each other where the new material is in the center, and the old material is at the outside of the cell.
A photograph of Christine Jacobs-Wagner smiling at the camera. She has short brown hair and green eyes.
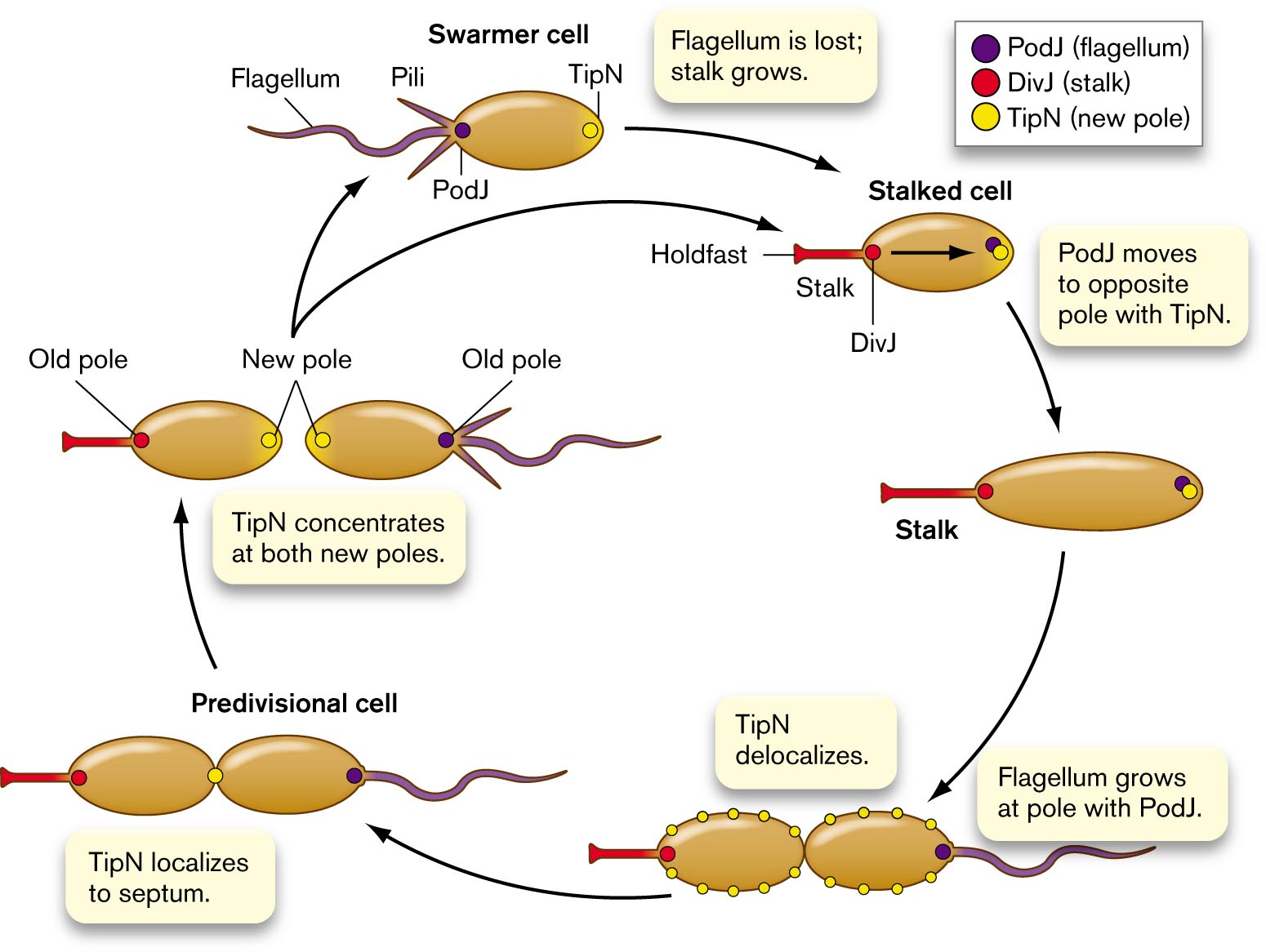
Cell development involves many such proteins working together. Figure 3.32 shows how TipN interacts with two other polar proteins: the flagellar marker PodJ, and the stalk marker DivJ. Each young cell (swarmer cell at top of cycle) has a new pole containing TipN. To prepare for cell division, the swarmer loses its flagellum. The flagellar pole then grows a stalk, with a holdfast for the cell to attach in a favorable environment. PodJ now migrates from the stalk pole to the opposite pole with TipN.
More information
An illustration of the Caulobacter cell cycle. The cycle begins from a swarmer cell, which is oval and has a flagellum and pili on one end of the cell. The P o d J protein is at the end of the cell by the flagellum and pili. The T i p N protein is at the opposite end of the cell. The flagellum is lost, and a stalk with a holdfast grows in its place leads to the stalked cell, P o d J has moved to the pole opposite the stalk with T i p N. D i v J is at the base of the stalk. The holdfast is at the end of the stalk. The stalked cell elongates. Then T i p N delocalizes around the membrane of the cell. A flagellum grows at the pole with P o d J. In the divisional cell, T i p N localizes to the septum, the connection between the two ovals. After the ovals separate into two cells, T i p N concentrates at both new poles. The old poles are where the stalk is on one cell and where the flagellum is on the other cell. The cell with the flagellum is a swarmer cell that restarts the cycle.
FIGURE 3.32 ■Cell cycle of Caulobacter. A swarmer cell loses its flagellum and grows a stalk. PodJ protein (purple) is at the flagellar pole, while DivJ protein (red) is at the stalk. TipN (yellow) is found at “new” poles (newly septated). TipN delocalizes and then localizes at the cell equator, midway between poles. The pole with PodJ grows a flagellum. The cell septates, forming two new poles, each containing TipN. The stalked cell still has DivJ at the stalk, and the new swarmer cell has PodJ at the flagellum.
Source: Modified from Melanie Lawler and Yves Brun. 2006. Cell124:891.
The stalk marker DivJ is now produced at the stalk pole, where PodJ was previously. As the stalked cell grows, TipN proteins delocalize around the cell. Then TipN localizes again at the middle, where the cell septates and divides. Once division is complete, TipN is concentrated at both new poles. The pole containing PodJ now grows a flagellum. The new flagellated cell is ready to find a new favorable environment to form a stalk and settle. Overall, throughout the cycle, a series of polar proteins localize and delocalize to define the polar functions.
Thought Question
3.10Figure 3.31 presents data from an experiment that allows the function of the TipN protein of Caulobacter to be visualized by microscopy. Can you propose an experiment with mutant strains of Caulobacter to test the hypothesis that one of the proteins shown in Figure 3.32 is required for one of the cell changes shown?
ANSWER ANSWER
The diagram of Figure 3.32 proposes that PodJ protein is required for a pole to develop a flagellum. Suppose we construct a mutant strain with a deletion of the gene podJ. This podJ mutant fails to express PodJ protein. When the podJ mutant is supplied with nutrients, the stalked cells should grow and fission, but the progeny from the plain pole should fail to grow a flagellum. The stalked progeny will continue to divide, producing a stalked cell and a cell with plain poles, lacking flagellum or stalk. Other results are possible, but the result described would be consistent with a requirement of PodJ for flagellar development.
Growth Asymmetry and Polar Aging
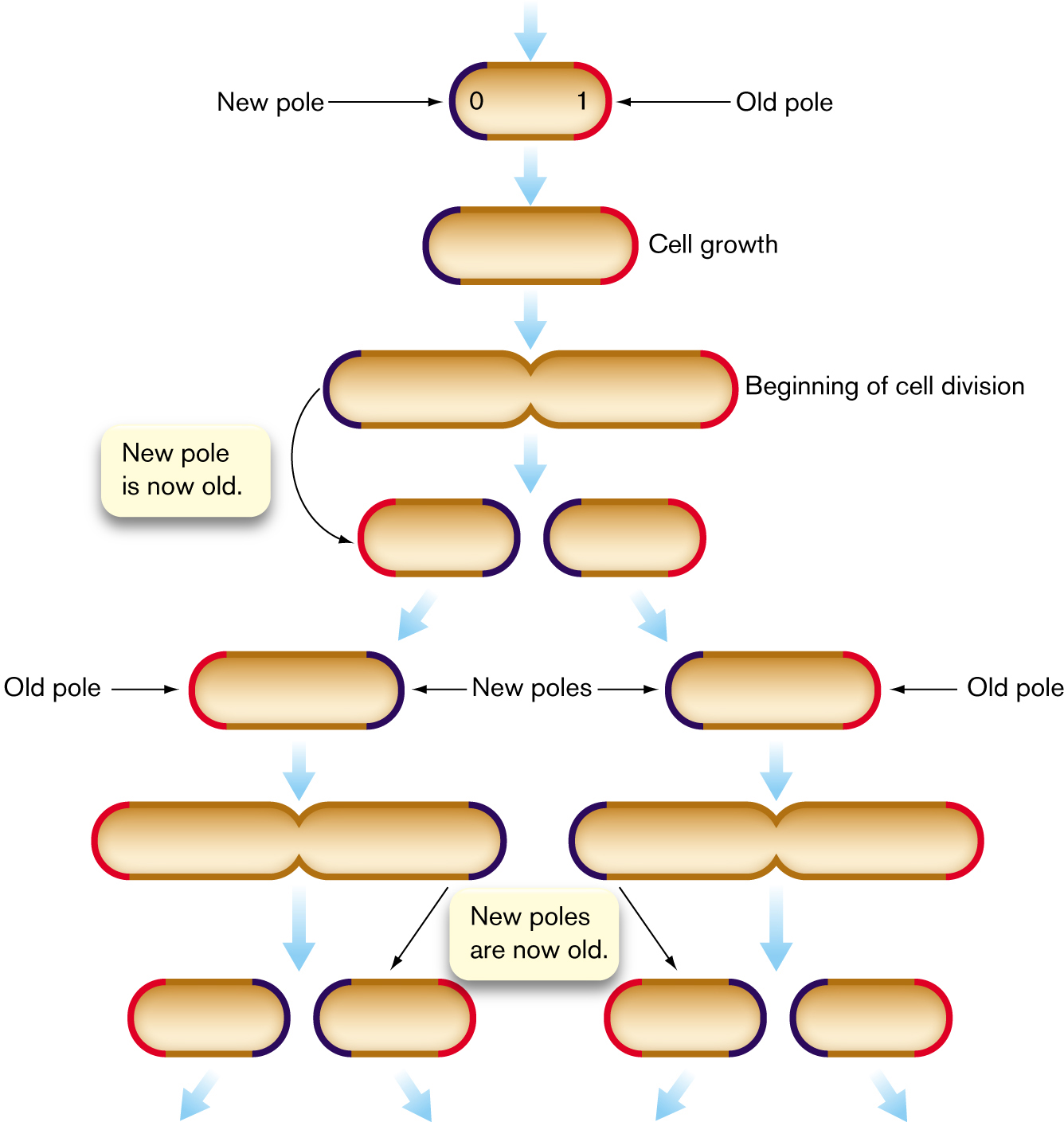
Does an apparently symmetrical cell such as E. coli actually possess two different polar forms? In fact, cell division generates daughter cells with chemically different poles (Fig. 3.33). Each cell starts out with one “old” pole (red in the figure) and one “new” pole (blue) where the parental cell septated. As the next cell divides, two daughter cells form, each with another “new” pole. But meanwhile, the “old” poles continue to age. With each generation, the polar cell wall material degrades slightly, increasing the chance of cell lysis. In a population of E. coli under environmental stress, at each cell division some members of the population die—of polar old age.
More information
An illustration shows the process of the generation of bacterial cells with an old pole and a new pole. The process of the cell begins with a new pole labeled 0 on the left, and an old pole labeled 1 on the right leads to cell growth, elongating the oval. At the beginning of cell division, the cell is pinched in the middle as two ovals prepare to separate into cells. A note reads a new pole is now old. So the left cell has an old pole on the left and a new pole on the right. The right cell has a new pole on the left and an old pole on the right. Each of these two cells undergoes cell division. One dividing cell results in two offspring, one of which has an old pole on the left and the other has a new pole on the left and an old pole on the right. The pole that was new on the right of the parent cell has become an old pole on the right side of the right offspring.
FIGURE 3.33 ■Bacterial cell division generates cells with an old pole and a new pole. Succeeding generations have cells with diverse combinations of new poles (blue), old poles (red), and very old poles (two or more generations, also red).
Source: Modified from Eric Stewart et al. 2005. PLoS Biol.3:e45
The cause of polar aging in stressed E. coli is the preferential accumulation of protein aggregates, which are nonfunctional and cannot be unfolded or degraded. For unknown reasons, proteins aggregate more frequently under a stressful condition, such as low pH or the presence of an antibiotic. Proteins damaged by a stressful condition are packed away in the cell’s older pole, allowing the new-pole cells to remain intact and grow faster. This asymmetrical cell provisioning may represent a form of “altruism” in which the older half cell promotes faster growth of the younger half cell.
Yet other kinds of cells grow by extending one pole only. The actinomycete Corynebacterium glutamicum, a soil bacterium useful for industrial production, positions its replisome at one cell pole. As DNA replication begins, a second replisome moves to the opposite pole, while new cell wall forms at the poles. In the next generation, the opposite pole possesses the replisome and undergoes extension. Unequal or unipolar cell extension is common among actinomycetes and mycobacteria, such as Mycobacterium tuberculosis.
Why does polar aging matter? One consequence of polar aging is that cells of different polar ages may differ in their resistance to antibiotics. This phenomenon could cause problems for antibiotic therapy. In M. tuberculosis, alternate polar aging generates variable resistance to antibiotics. The result may give tuberculosis bacteria the opportunity to “try out” resistance to various antibiotics applied in chemotherapy.
An extreme form of asymmetrical growth is endospore formation by Firmicutes such as Bacillus and Clostridium species. An endospore is an inert but viable cell form, having no active metabolism but capable of germination under the right conditions. Under starvation, desiccation, or other stress conditions, a bacterium can undergo an asymmetrical cell division to develop an endospore at one end. Endospore formation requires an extreme form of cellular altruism, in which the mother cell sacrifices itself for the spore-forming cell. The process generates an endospore capable of remaining dormant but viable for thousands of years.
Note: Endospore formation is covered in detail in Section 4.6.
Membrane Vesicles
Our concept of the cell assumes a defined boundary of membrane that encloses the cell’s contents and separates them from the external space. The cell’s cytoplasm is a precious limited resource. Yet surprisingly, isolated microbial cells continually export bits of cytoplasm in membrane vesicles. Some kinds of microbes share their materials with other cells—even cells of other species—via intercellular nanotubes. How does this cytoplasmic sharing serve the cell?
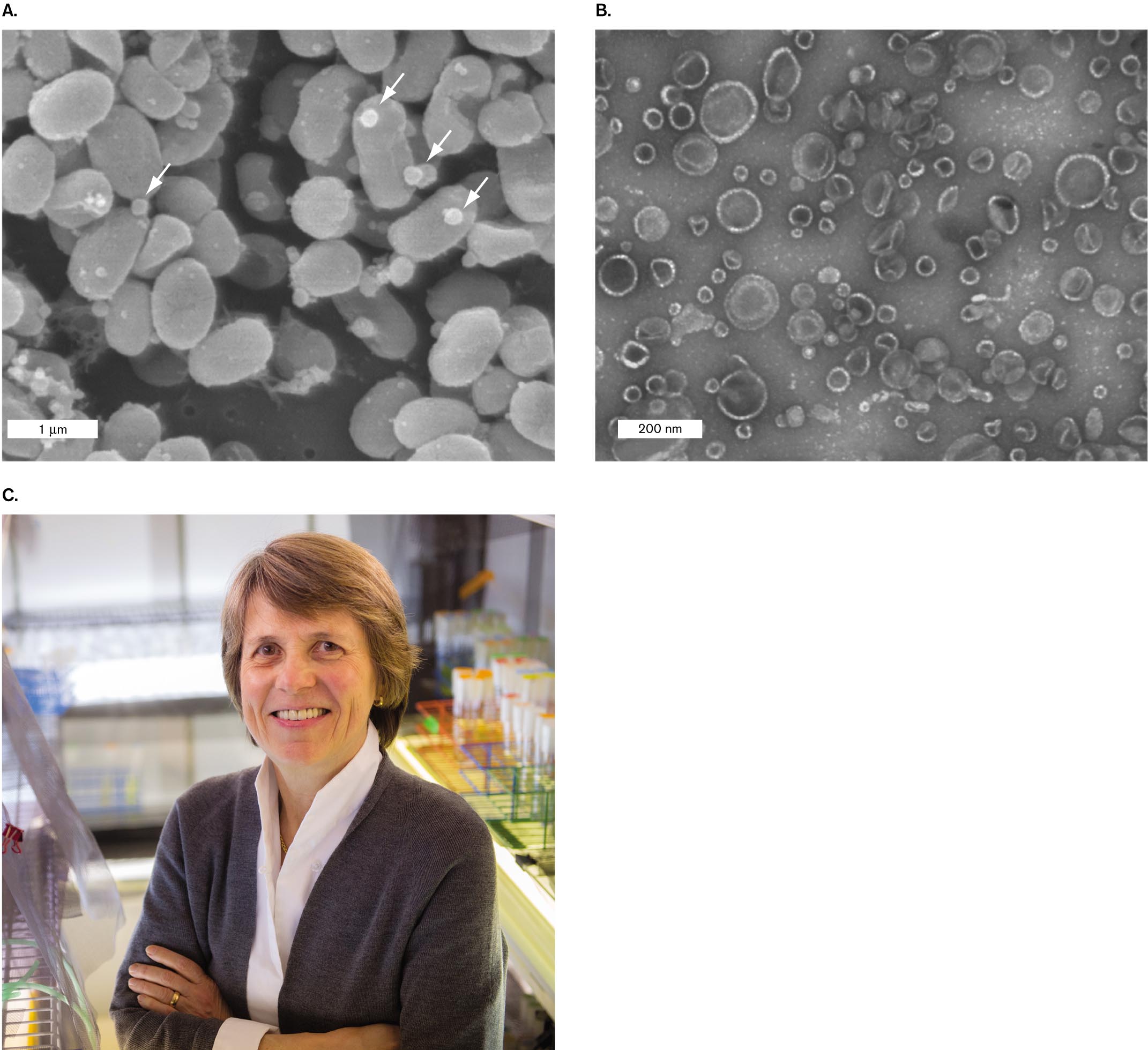
Membrane vesicles carry proteins and nucleic acids. An example of cytoplasmic export via membrane vesicles in the marine cyanobacterium Prochlorococcus was documented by Sallie Chisholm at the Massachusetts Institute of Technology. Prochlorococcus is one of the smallest yet most abundant phototrophs in Earth’s oceans, having a global population estimated at 3 × 1027 cells, which perform 20% of all photosynthesis in our oceans. Because Prochlorococcus cells are so small and their nutrients so scarce, it is remarkable that these tiny cells release their cytoplasm by pinching off vesicles (Fig. 3.34). Chisholm and her postdoctoral fellow analyzed these vesicles by ultracentrifugation (discussed in Section 3.1) and biochemical assays of their contents. The vesicles were found to contain diverse proteins, RNA molecules, and even fragments of DNA.
More information
Two micrographs of Prochlorococcus cyanobacteria and a photograph of Sallie Chisholm.
A scanning electron micrograph of Prochlorococcus cyanobacteria releasing vesicles of the cell membrane. The image is packed with bacteria. The bacteria are circular, each with a diameter of about 1 micrometer. Small circular vesicles are visible and are floating freely.
A transmission electron micrograph of Prochlorococcus vesicles. The vesicles appear as transparent circular shapes less than 200 nanometers in diameter.
A photograph of Sallie Chisholm smiling at the camera. She has short blonde hair and brown eyes. She is standing in a laboratory.
FIGURE 3.34 ■Marine bacteria release membrane vesicles.A.Prochlorococcus cyanobacteria release vesicles (arrows) of cell membrane into the open ocean (SEM). B. Vesicles collected from Prochlorococcus (TEM). C. Sallie Chisholm, professor of environmental studies, pioneered the study of Prochlorococcus and its significance for marine ecology. S. J. BILLER ET AL. 2014. SCIENCE343 (6167):183–186, FIG. 1AS. J. BILLER ET AL. 2014. SCIENCE343 (6167):183–186, FIG. 1BRICHARD HOWARD
What functions are served by vesicle production that outweigh the loss of precious resources? Chisholm finds evidence for several possibilities:
Attraction of partner heterotrophs. Heterotrophic bacteria attracted by released carbon sources consume the excess oxygen and reactive oxygen species (ROS) produced by cyanobacterial photosynthesis. Cyanobacteria require heterotrophic partners for optimal growth.
Phage decoys. Bacteriophages readily infect Prochlorococcus and deplete its populations. But the bacterial membrane vesicles possess envelope receptors for phages, which can trap the phages and prevent them from infecting cells.
DNA transfer. The DNA released by Prochlorococcus may provide useful genetic traits for other members of the population as a form of horizontal gene transfer.
Another system in which membrane vesicles are shared is that of human gut bacteria such as Bacteroides and related anaerobes. One way the gut environment differs from the open ocean is in the abundance of nutrients available. In the gut, many anaerobes release vesicles of partly digested complex polysaccharides for further catabolism by other species. Often, both community members benefit as a result. Gut microbial interactions are discussed further in Chapter 21.
Membrane Extensions and Nanotubes
From the observations of the early microscopists of the nineteenth century, most bacteria were thought to have relatively simple shapes such as rods or spheres. But cryo-electron microscopy revealed surprisingly delicate extensions of the cell or outer membranes, such as filaments and “pearling” chains of vesicles. What is the function of these cell extensions? In some cases, cell extensions expand the cell’s reach for scarce nutrients (see Chapters 4 and 18). The purpose of other membrane extensions remains unclear, but exciting discovery continues.
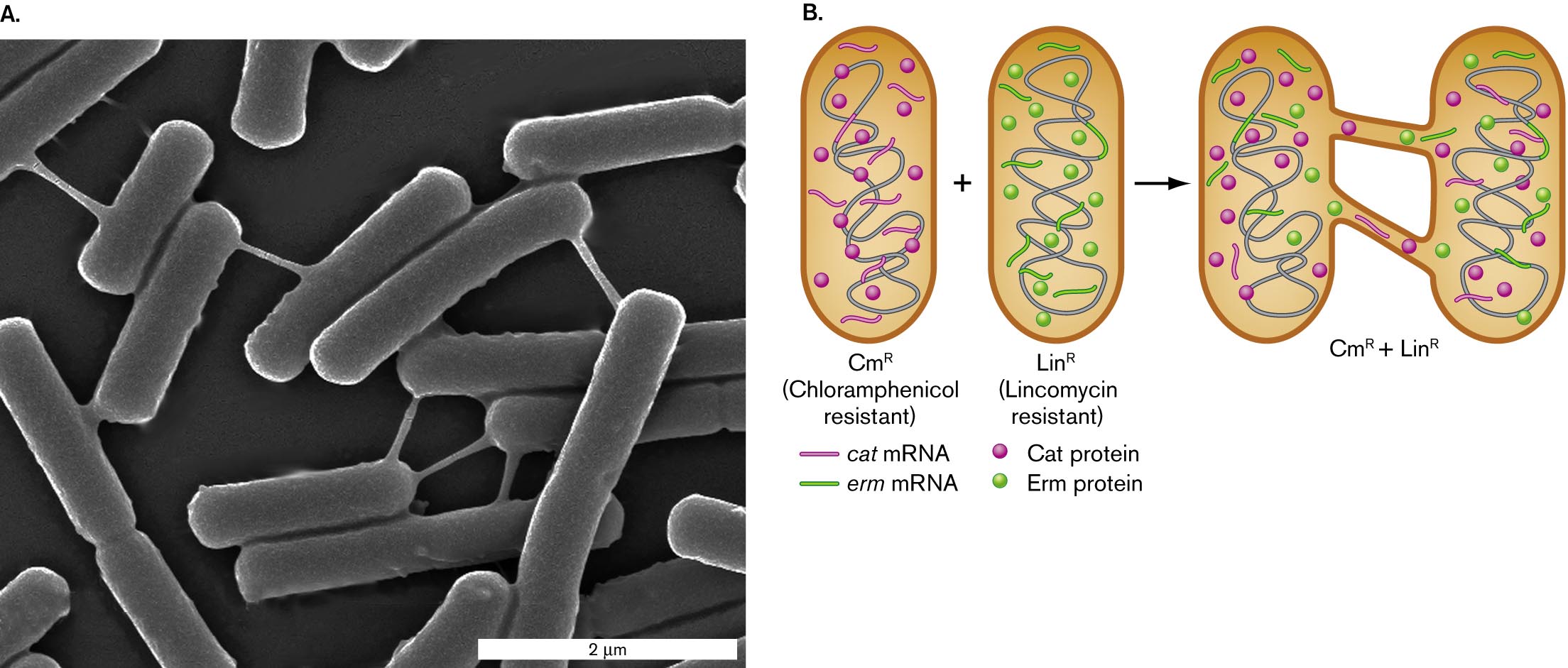
Some bacteria and archaea can form membrane extensions that merge directly with the membranes of neighboring cells. Sigal Ben-Yehuda and students at the Hebrew University of Jerusalem revealed such extensions, called intercellular nanotubes, between cells of Bacillus subtilis (Fig. 3.35). B. subtilis is a Gram-positive bacterium common in soil, a highly complex environment full of diverse nutrients and antimicrobial toxins (discussed in Chapter 21). The nanotubes enable bacteria to directly share proteins and messenger RNA that encodes products useful under hostile conditions, such as exposure to antibiotics (Fig. 3.35B). Ben-Yehuda showed that two Bacillus cells encoding resistance to two different antibiotics—chloramphenicol (Cat protein) and lincomycin (Erm protein)—could share their resistance proteins and messenger RNA via nanotubes. The connected bacteria resist both antibiotics.
More information
A micrograph and an illustration of nanotubes connecting cells of Bacillus subtilis.
A micrograph of Bacillus subtilis cells connected by nanotubes. Several rod shaped bacteria are visible. The bacteria are about 2 micrometers long and 0.5 micrometer wide. The bacteria are connected by thin tubes.
An illustration of nanotubes between cells. A rod shaped bacterium is shown, labeled C m superscript R, or chloramphenicol resistant. This bacterium contains several strands of c a t m R N A and several C a t proteins. Another rod shaped bacterium is shown, labeled L i n superscript R, or Lincomycin resistant. This bacterium contains several strands of e r m m R N A and several E r m proteins. When nanotubes form between the two bacteria, the resistance genes and proteins are shared between the two.
FIGURE 3.35 ■Intercellular nanotubes.A.Bacillus subtilis bacteria connected by intercellular nanotubes, which pass material from one cell to the next. B. Nanotubes connect bacteria with two different genes encoding proteins that confer resistance to an antibiotic: chloramphenicol (Cat) or lincomycin (Erm). The connected bacteria share mRNA and resistance proteins for both chloramphenicol and lincomycin resistance. CmR = chloramphenicol resistance; LinR = lincomycin resistance. G. P. DUBEY AND S. BEN-YEHUDA. 2011. CELL144:590
A similar experiment showed that even bacteria of different species can share beneficial components of cytoplasm. Christian Kost and students at Max Planck Institute for Chemical Ecology in Jena, Germany, used fluorescence microscopy to show that Escherichia coli bacteria can form nanotubes with the Gram-negative bacterium Acinetobacter baylyi. The nanotubes facilitate exchange of different amino acids between these two species. Remarkably, the nanotubes form only when the two types of cells each produce an amino acid lacking in the other. Thus, nanotubes facilitate metabolic cross-feeding.
Archaea show various kinds of intercellular nanotubes that are essential parts of the cell. Examples are found in Pyrodictium and Thermococcus species, as well as the Asgard archaeon, Prometheoarchaeum synthrophicum (see Chapter 1 opening image). For more examples, see Chapter 19.
Note: Bacterial nanotubes between cells remain a subject of controversy, because some laboratories fail to replicate their discovery, even using the same bacterial strains. In Archaea, intercellular nanotubes are more clearly documented.
Thought Question
3.11 Could two bacteria share protein complexes via nanotubes? What about hydrogen molecules (H2) as electron donors?
ANSWER ANSWER
Bacterial cells share proteins via nanotubes, such as enzymes for carbohydrate catabolism. In principle, a nanotube could be wide enough to allow transmission of ribosomes. However, nanotubes could not share dihydrogen molecules, because H2 is a gas that penetrates membranes and would escape through the nanotube walls.
SPECIAL TOPIC 3Bacteria Reach Out with Pearling Tubes and Nanopods
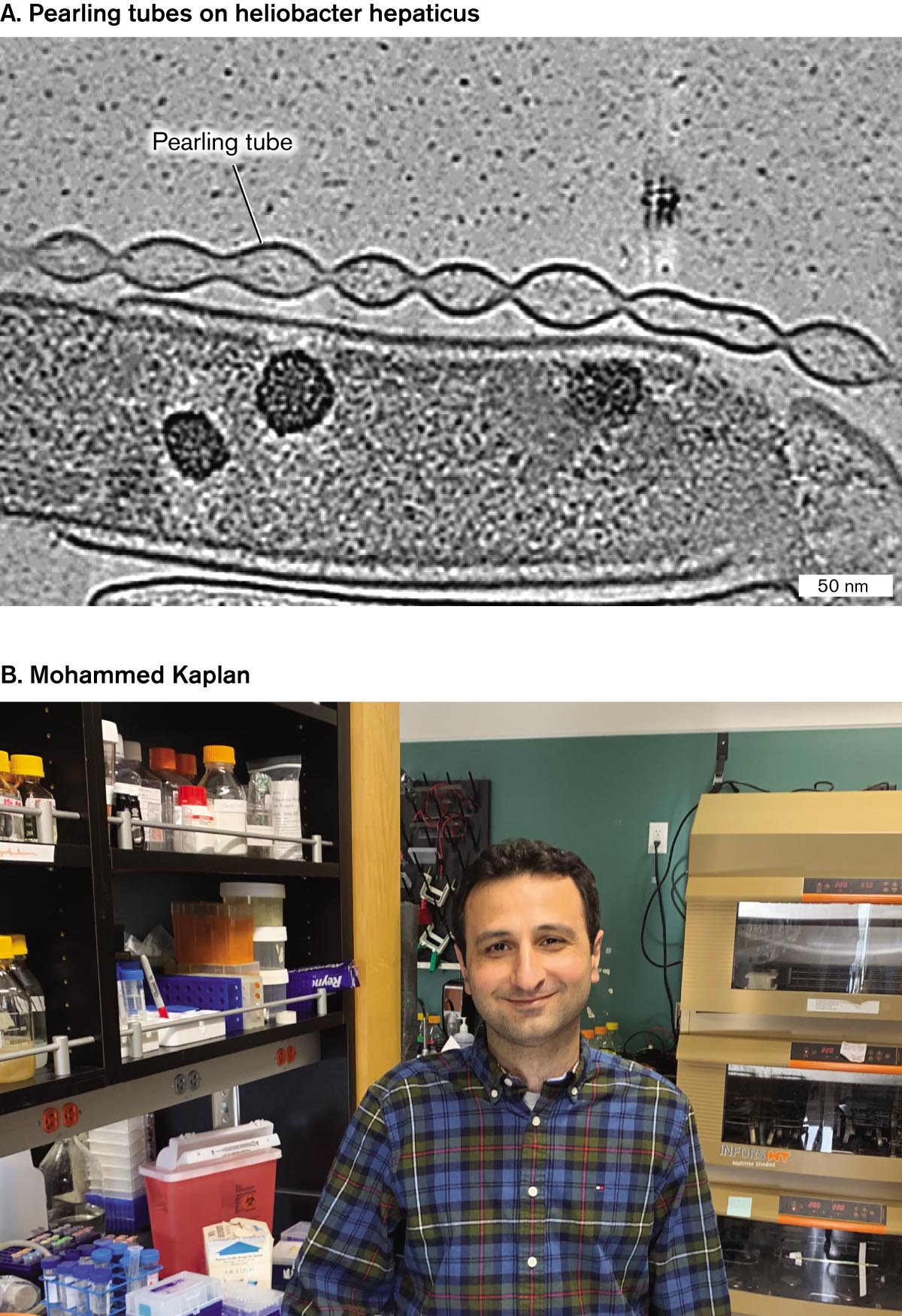
What if you found an intricate membrane structure like a string of pearls and had no idea what it does? Mohammed Kaplan, along with colleagues at the California Institute of Technology and several other universities, hunted for novel membrane extensions in cryo-electron tomograms (tomography slices) from 90 species of bacteria. The resolution and clarity of cryo-EM and the absence of heavy-metal stains revealed fine structures that were missed by earlier electron microscopy. Thirteen of the bacteria showed novel structures never before reported—and whose functions remain unclear (Fig. ST 3.1).
More information
A micrograph of pearling tubes on Helicobacter hepaticus and a photo of Mohammed Kaplan.
A cryo-electron micrograph of pearling tubes on Helicobacter hepaticus. A rod shaped cell is partially visible and appears nearly transparent. The cell is about 100 nanometers wide and more than 400 nanometers long. A structure rests on the one edge of the cell. The structure appears as a chain of pearls and is labeled as a pearling tube.
A photo of Mohammed Kaplan in a laboratory. He has dark brown hair and brown eyes. He is wearing a plaid shirt and sitting at a lab bench. He is smiling at the camera.
FIGURE ST 3.1 ■Mysterious membrane extensions.A. Pearling tubes of Helicobacter hepaticus (cryo-EM). B. Mohammed Kaplan, graduate student in the lab of Grant Jensen, California Institute of Technology.
Source: Mohammed Kaplan et al. 2021. Elife10:e73099.
M. KAPLAN ET AL. 2021. ELIFE. 10:E73099MOHAMMED KAPLAN
Helicobacter hepaticus is a spiral-shaped Gram-negative proteobacterium that infects the livers of mice and is a relative of the ulcer-causing human pathogen Helicobacter pylori. The tomograms of H. hepaticus showed long strings of interconnected vesicles of outer membrane, each with a round shape like pearls on a string (Fig. ST 3.1A). The function of these vesicles is not known, but similar “pearling tubes” are found in aquatic Shewanella bacteria, where they may carry electricity. Bacterial electricity and its use for fuel cells are described in Chapter 14.
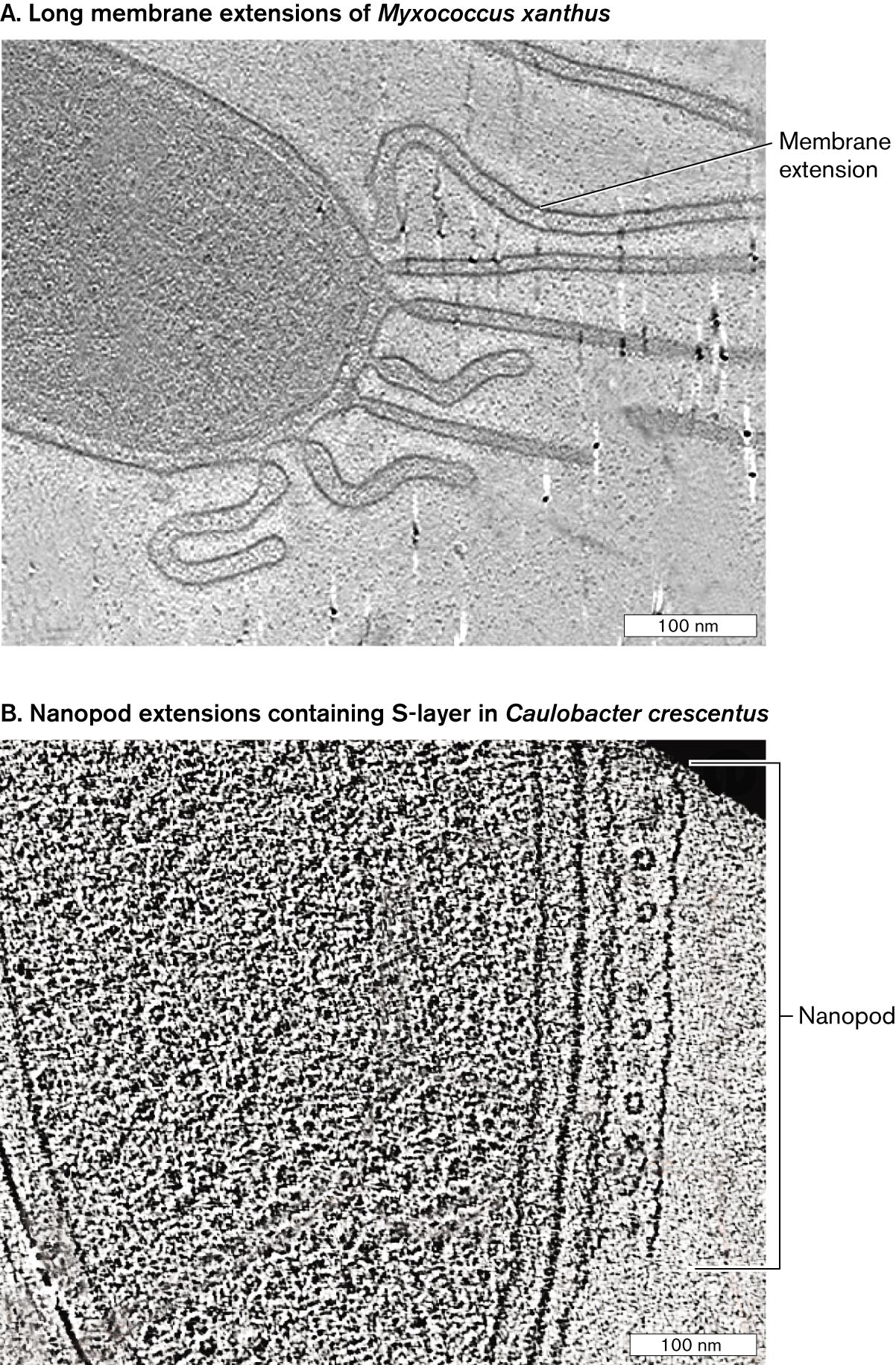
A different kind of outer membrane extension was found in Myxococcus xanthus, a predatory bacterium that collects in swarms to form a fruiting body and spores. M. xanthus cells extend multiple long tubes of outer membrane (Fig. ST 3.2A). The function of these tubes is unknown, although in other microbes lengthy extensions help the cell obtain nutrients from nutrient-poor environments. Similar membrane extensions are found in Caulobacter crescentus, but its tubes are filled with S-layer subunits. The subunits are wrapped in membrane, like a sandwich in plastic wrap. These S-layer tubes are called nanopods (Fig. ST 3.2B). Because S-layer proteins generally protect a cell surface, it is puzzling to find them contained by a membrane. We do not know what substances are contained by pearling tubes, long extensions, or nanopods. We only know that the more we investigate microbial cells, the more amazing and unexpected components we find.
More information
Two cryo-electron micrographs of membrane extensions in different bacterial species.
A cryo-electron micrograph of long membrane extensions in Myxococcus xanthus. A small portion of a rod shaped cell is visible. Long tube like tendrils extend from the end of the cell. The tendrils are labeled membrane extensions. The tendrils are longer than 100 nanometers.
A cryo-electron micrograph of nanopod extensions in Caulobacter crescentus. A small portion of a rod shaped cell is visible. Along the outer edge of one side of the cell is a tube like structure with several circular components evenly space within the tube. The structure is parallel to the long edge of the cell and about 300 nanometers long. The structure is labeled nanopod.
FIGURE ST 3.2 ■Membrane extensions and nanopods.A.Myxococcus xanthus membrane extensions (cryo-EM). B.Caulobacter crescentus nanopod (cryo-EM). Source: Mohammed Kaplan et al. 2021. Elife10:e73099.M. KAPLAN ET AL. 2021. ELIFE. 10:E73099M. KAPLAN ET AL. 2021. ELIFE. 10:E73099
RESEARCH QUESTION
What experiments might you perform to reveal the function of a mysterious bacterial structure?
Kaplan, Mohammed, Georges Chreifi, Lauren A. Metskas, Janine Liedtke, Cecily R. Wood, et al. 2021. In situ imaging of bacterial membrane projections and associated protein complexes using electron cryo-tomography. Elife10:e73099.
To Summarize
The poles of a bacterial cell may differ in form and function.Caulobacter crescentus has one plain pole and one pole that has either a flagellum or a stalk. A stalked cell fissions to produce one stalked cell and one flagellar cell.
The two bacterial poles differ in age. One pole arises from the septum of the parental cell, whereas the other pole arises from a parental pole. In E. coli, successive cell divisions yield progeny with a mixture of polar ages. Cells with a very old pole may cease replication and die.
Polar aging is increased by stress. Environmental stress, such as an antibiotic or low pH, causes protein aggregates to collect at the cell’s older pole. Actinobacterial cells extend at alternating poles.
Membrane vesicles transmit cytoplasmic contents. Vesicles share proteins, nucleic acids, and other cytoplasmic contents with the exterior environment and other cells.
Intercellular nanotubes directly share cytoplasmic contents. Nanotubes between individual bacteria share drug resistance, cross-feed nutrients, and mediate electron transfer.
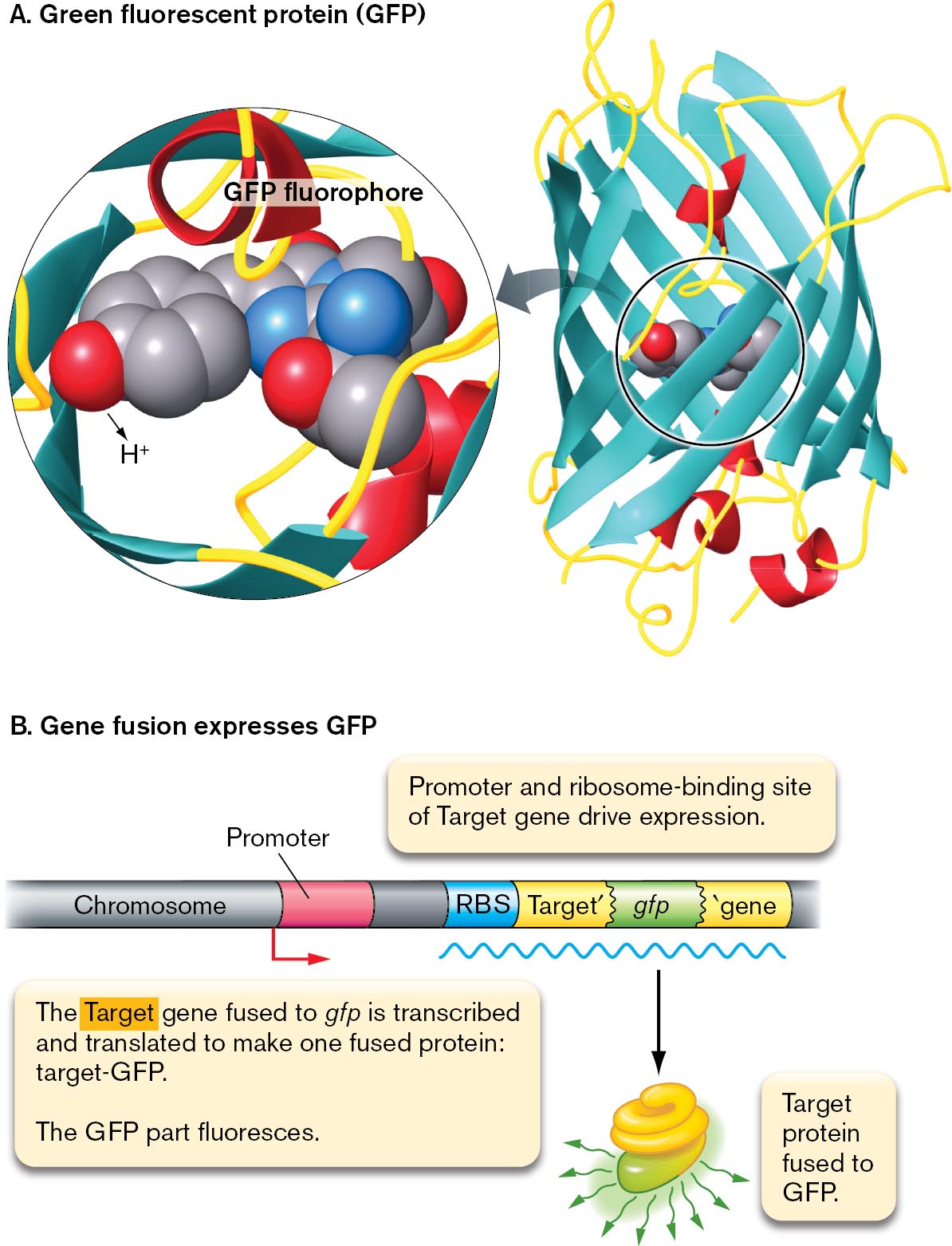
A model of G F P fluorophore and a model of G F P gene fusion.
A space filling model and the protein structure of G F P fluorophore are shown. The protein structure of G F P shows the fluorophore within barrel-shaped beta-sheets. A magnified view of the fluorophore shows a space-filling model of the three amino acids serine, tyrosine, and glycine, bonded.
A model of G F P gene fusion. A chromosome sequence has the promoter, R B S, target gene prime, g f p, and prime gene. An arrow points forward from the promoter, and an m R N A strand lies below the chromosome. From the m R N A strand, an arrow points down to show a truncated protein fused with another protein fluorescing. The fused protein is labeled, target protein fused to G F P. Accompanying text reads, The target gene fused to g f p is transcribed and translated to make one fused protein: target hyphen G F P. The G F P part fluoresces. Above the chromosome sequence, text reads, promoter and ribosome binding site of target gene drive expression.
FIGURE 2.29 ■The fluorophore green fluorescent protein (GFP).A. Green fluorescent protein (GFP) is expressed endogenously by the cell. Blowup: Three GFP amino acid residues (serine, tyrosine, and glycine) condense to form the fluorophore. B. The gene encoding GFP can be fused to a target gene (Target′-gfp). The fused gene then expresses a fused protein in which the GFP portion fluoresces. The fluorescent protein is expressed under control of the target gene promoter and ribosome-binding site (RBS).
 ANSWER
ANSWER ANSWER
ANSWER