All three domains of life—Bacteria, Archaea, and Eukarya—consist of one or more cells. Viruses are acellular, but they require a cell to infect. Certain features of a living cell are distinctive to one or more domains:
Bacteria and Archaea are prokaryotes (cells that lack a nucleus). Bacteria have phospholipid bilayers similar to those of eukaryotes, but archaea have unique membrane and envelope structures such as ether membranes that enable survival in extreme environments. Nevertheless, many kinds of archaea live in moderate environments, such as soil, water, or human skin.
Eukaryotic cells have extensive membranous organelles. Organelles such as the endoplasmic reticulum and Golgi complex are reviewed in eAppendix 2. The mitochondria and chloroplasts of eukaryotic cells evolved by endosymbiosis with engulfed bacteria (see Chapter 17). Diverse microbial eukaryotes, such as fungi and protists, are explored in Chapter 20.
Chapter 3 focuses primarily on bacteria. Most bacteria share these traits:
Thick, complex outer envelope. The envelope protects the cell from environmental stress and mediates exchange with the environment.
Compact genome. Prokaryotic genomes are compact, with relatively little noncoding DNA. Small genomes maximize the production of cells from limited resources.
Tightly coordinated functions. The cell’s parts work together in a highly coordinated mechanism, which may enable a high rate of reproduction.
Next, we embark on a tour of a typical Gram-negative bacterial cell. Along the way, we learn how our understanding of this model cell has emerged from microscopy, cell fractionation, and genetic analysis.
Note: This chapter assumes an understanding of introductory biology and chemistry. For review, see eAppendix 1 (“Biological Molecules”) and eAppendix 2 (“Introductory Cell Biology”).
Model of a Bacterial Cell
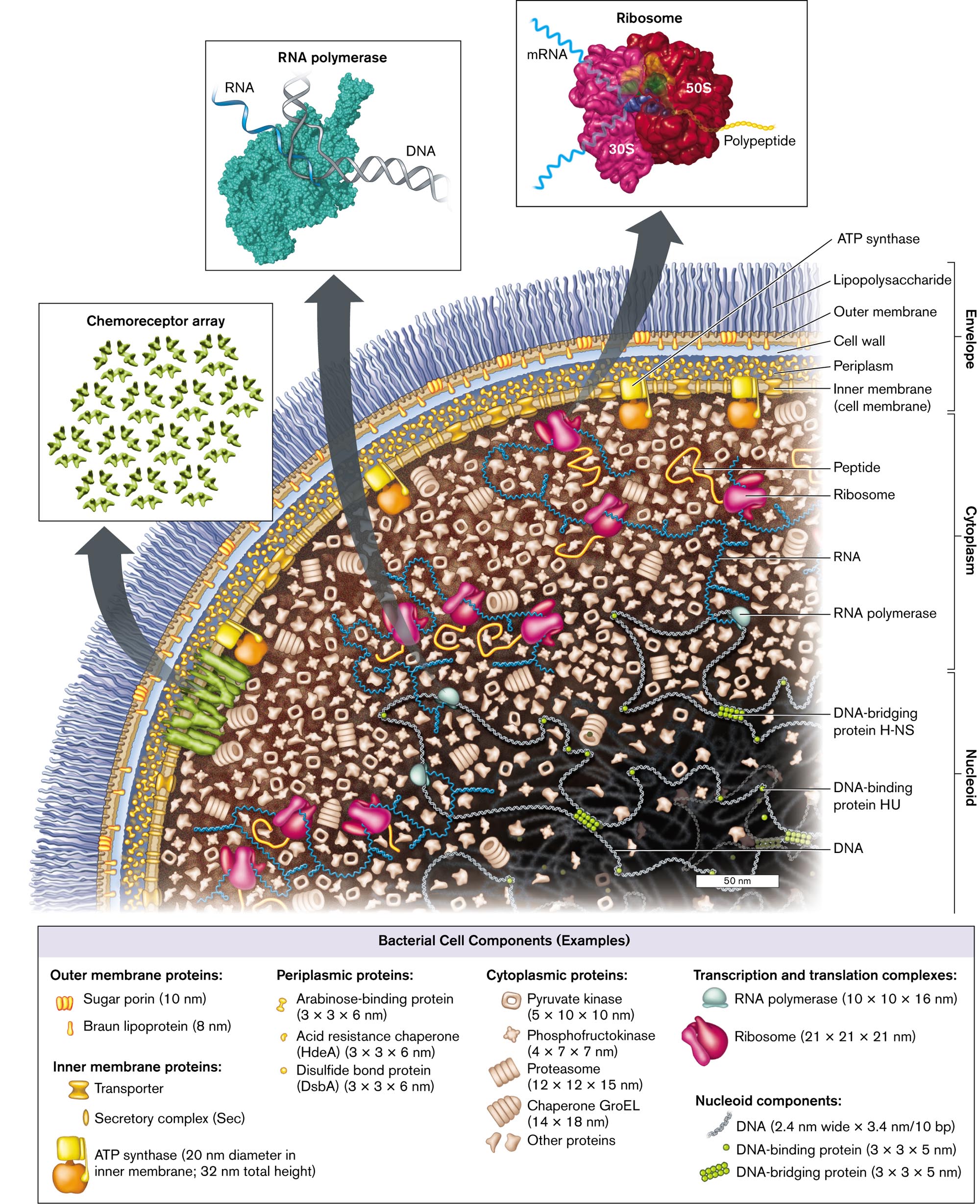
A cell is more than a “soup” full of ribosomes and enzymes. In fact, the cell’s parts fit together in a structure that is ordered, though flexible. Cryo-electron microscopy (cryo-EM; see Section 2.6) reveals distinct parts of a cell (see the Chapter 3 opening image). On the basis of many such microscope images, we offer a model of a typical bacterium (Fig. 3.1). This model represents Escherichia coli, a common Gram-negative bacterium that resides on your gut epithelium. Its general features apply to many kinds of bacteria, particularly the Gram-negative inhabitants of your colon such as Proteobacteria and Bacteroidetes. Remember that we cannot literally “see” molecules within a cell, but microscopy and subcellular analysis provide the evidence for our model.
More information
An illustration of the cell components and structure of Escherichia coli. The bacterium consists of small molecules such as nucleoid, cytoplasm, and envelope. The nucleoid contains D N A, D N A-binding protein H U, and D N A-bridging protein H-N S. The cytoplasm contains R N A polymerase, R N A, peptides, and ribosomes. The envelope contains the inner Membrane, or cell membrane, periplasm, the cell wall, the Outer Membrane, and lipopolysaccharides. A flagellar motor connects to the inner Membrane and travels outward. An inset shows a model of an R N A polymerase with the labeled parts D N A and R N A. Another inset from the bacterium shows a model of several D N A-binding proteins is labeled as Chemoreceptor array. The third inset shows a model of Ribosome bonded with m R N A, 30 S, 50 S, and polypeptide chain at a magnification of 50 nanometers. The fourth inset shows a microscopic view of the whole bacterium, and the labeled parts are the envelope, the nucleoid, and ribosomes at a magnification of 0.25 micrometers. A box containing examples of bacterial cell components is as follows: Outer membrane proteins: Sugar porin, 10 nanometers long, and Braun lipoprotein, 8 nanometers long; Inner membrane proteins: A T P synthase, 20 nanometers diameter in the inner Membrane; 32 nanometers total height, Secretory complex, abbreviated S e c, and Transporter. Periplasmic proteins: Arabinose-binding protein, dimensions 3 times 3 times 6 nanometers, Disulfide bond protein, abbreviated D s b A, dimensions 3 times 3 times 6 nanometers, Acid resistance chaperone, abbreviated H d e A, dimensions 3 times 3 times 6 nanometers. Cytoplasmic proteins: Pyruvate kinase, dimensions 5 times 10 times 10 nanometers, Phosphofructokinase, dimensions 4 times 7 times 7 nanometers, Proteasome, dimensions 12 times 12 times 15 nanometers, Chaperone G r o E L, dimensions 14 times 18 nanometers, Other proteins. Transcription and translation complexes: R N A polymerase, dimensions 10 times 10 times 16 nanometers, Ribosome dimensions 21 times 21 times 21 nanometers. Nucleoid components: D N A, dimensions 2.4 nanometers wide times 3.4 nanometers per 10 b p, D N A-binding protein, dimensions 3 times 3 times 5 nanometers, and D N A-bridging protein, dimensions 3 times 3 times 5 nanometers.
FIGURE 3.1 ■Escherichia coli: a Gram-negative bacterium of the gut microbiome. The envelope includes the outer membrane, the cell wall and periplasm, and the inner (cell) membrane with an embedded chemoreceptor array. The cytoplasmcontains enzymes, messenger RNA (mRNA) extending out of the nucleoid, and ribosomes. Ribosomes translate the mRNA to make proteins, which are folded by chaperones. The nucleoidcontains the chromosomal DNA wrapped around binding proteins. (PDB codes: ribosome, 1GIX, 1GIY; RNA polymerase, 1MSW)
Within a cell, the cytoplasm consists of a gel-like network composed of proteins and other macromolecules. The cytoplasm is contained by a cell membrane, or plasma membrane. For E. coli, a Gram-negative bacterium, the plasma membrane is called the inner membrane, in order to distinguish it from the additional outer membrane. The inner membrane is composed of phospholipids, transporter proteins, and other molecules. This membrane prevents cytoplasmic proteins from leaking out and maintains gradients of ions and nutrients.
Between the inner and outer membranes lies the cell wall, a fortress-like structure composed of sugar chains linked covalently by peptides (peptidoglycan). The cell wall forms a single molecule that surrounds the cell. A Gram-positive species would have the cell wall outside its one plasma membrane. The wall material is flexible, but it limits expansion of the cytoplasm, keeping the cell membrane intact when water flows in. Like air inside a balloon, the resulting turgor pressure (expansion force) makes the cell rigid.
In a Gram-negative bacterium, such as our model E. coli (Fig. 3.1), the cell wall lies within the periplasm, a water-filled space containing nutrient-binding proteins and secretion machines. Outside the cell wall lies the outer membrane of phospholipids and lipopolysaccharides (LPS), a class of lipids attached to long polysaccharides (sugar chains). The LPS layer may be surrounded by a thick capsule. The capsule polysaccharides form a slippery layer that inhibits phagocytosis by amebas or white blood cells. The inner membrane, cell wall, and outer membrane constitute the Gram-negative cell envelope.
The bacterial envelope includes cell-surface proteins that enable the bacterium to interact with specific host organisms. For example, E. coli cell-surface proteins help the bacterium colonize the human intestinal epithelium. The nitrogen-fixing symbiont Sinorhizobium has cell-surface proteins that help the bacteria colonize legume plants for nitrogen fixation.
To sense the environment and to direct swimming, motile bacteria possess an array of chemoreceptors (receptor array), usually situated at one pole (rounded end of a rod-shaped cell). The receptors bind molecules from outside the cell or the periplasm and convert this binding information into signals within the cytoplasm. The signaling molecules direct the rotation of flagella (singular, flagellum), propeller-like organelles for swimming, described in Section 3.6. The amazing phenomenon of chemotaxis, how bacteria choose which direction to swim, is explored later in Chapter 12.
The Membrane Is a Two-Dimensional Fluid of Lipids and Proteins
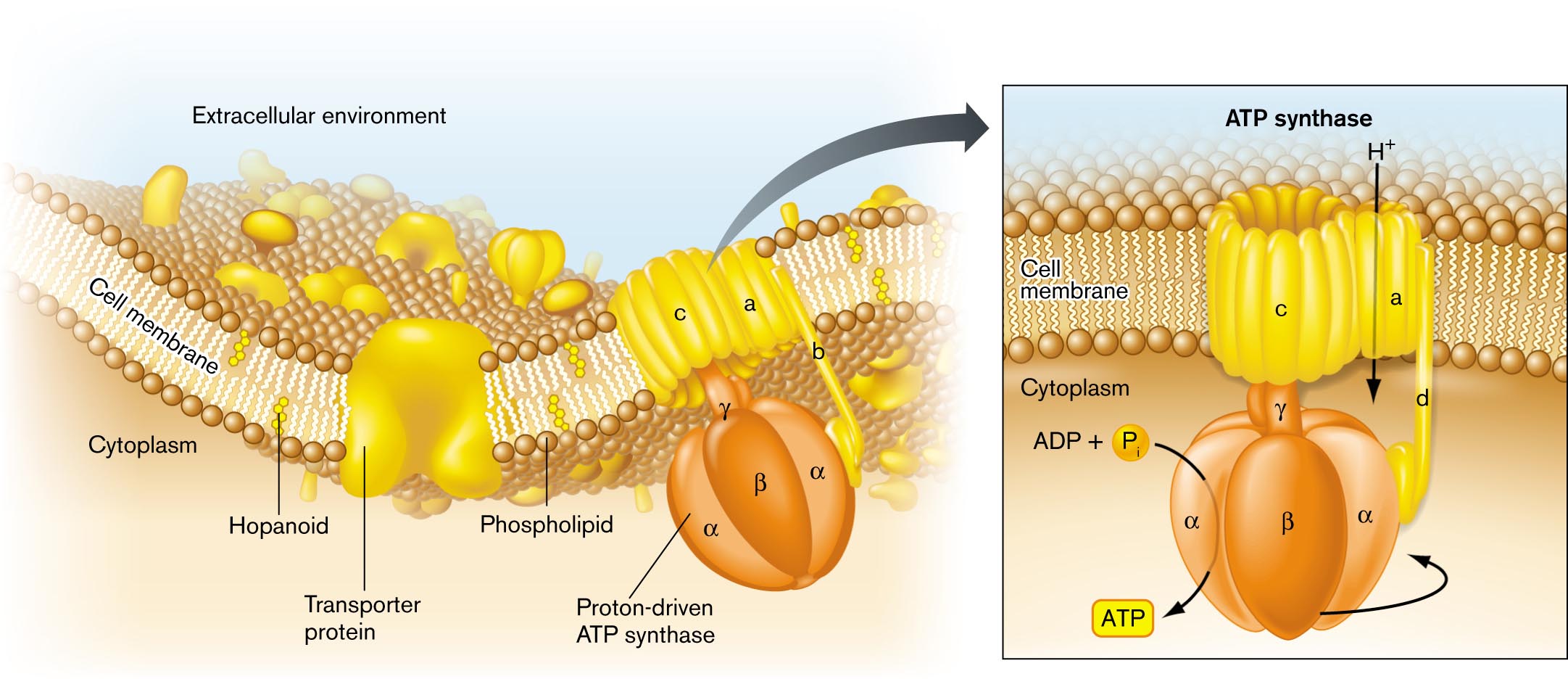
The cell membrane (or inner membrane, for a Gram-negative cell) contains the cytoplasm within the external medium, and it can maintain aqueous compartments with very different properties. For example, the concentration of hydrogen ions (H+) can be a hundred times greater outside the cell than inside. The membrane consists of a phospholipid bilayer (double layer of phospholipids) including lipid-soluble proteins. The bilayer behaves as a two-dimensional fluid, within which proteins and lipids can diffuse (Fig. 3.2). The proteins form about half the mass of the membrane and provide specific functions, such as nutrient transport. We will examine this structure in detail in Section 3.2.
More information
An illustration of the general structure of a bacterial cell membrane. The area beyond the cell membrane is labeled the extracellular environment. The area within the cell is labeled cytoplasm. The cell membrane is comprised of a phospholipid bilayer. Several hopanoids are layered within the membrane. A transporter protein embedded in the membrane opens to both the extracellular environment and the intracellular environment. A proton-driven A T P synthase is embedded in the membrane and opens to the both the extracellular environment and the intracellular environment. Further detail on the A T P synthase describes that hydrogen is taken into the cell. The proton driven pump is located in the cell interior, and uses A D P plus inorganic phosphate to generate A T P.
FIGURE 3.2 ■Bacterial cell membrane. The cell membrane consists of a phospholipid bilayer, in which the hydrophobic fatty acid chains are directed inward, away from water. The bilayer contains stiffening agents such as hopanoids. Half the membrane volume consists of proteins such as the ATP synthase protein complex.
Proteins embedded in the cell membrane often function together as a “complex.” The subunits of a complex fit together closely, like puzzle pieces. A famous protein complex is ATP synthase, an enzyme that couples the flow of H+ ions across the membrane to the synthesis of ATP. The flow of H+ ions through the ATP synthase is driven by the charge difference (more negative inside the cell) and the concentration difference (lower H+ concentration inside). Thus, a surprisingly intricate “molecular machine” can sit in the membrane and synthesize ATP for the cell. Energy transduction by ion gradients is discussed further in Chapter 4 and Chapter 14.
Within the cell, the cell membrane and envelope provide an attachment point for one or more chromosomes. The chromosome is organized within the cytoplasm as a system of looped coils called the nucleoid. Unlike the round, compact nucleus of eukaryotic cells, the bacterial nucleoid is not enclosed by a membrane. Instead, loops of DNA extend throughout the cytoplasm. The DNA is transcribed by RNA polymerase to form messenger RNA (mRNA), as well as transfer RNA (tRNA) and ribosomal RNA (rRNA). As the mRNA transcripts extend, they bind ribosomes to start synthesizing polypeptide chains. As the polypeptides grow, they fold into their three-dimensional shape. This process may require help from protein complexes called chaperones. The concept of information flow from DNA to RNA to protein is presented in Chapters 7–10.
Biochemical Composition of Bacteria
The bacterial cell model in Figure 3.1 represents the shape and size of cell parts but tells us little about the chemistry of the cell or about the cell’s environment. Chemistry explains, for example, why wiping a surface with ethanol kills microbes, whereas water has little effect. Water is a universal constituent of cytoplasm but is excluded by cell membranes. Ethanol, however, dissolves both polar and nonpolar substances; thus, ethanol disintegrates membranes and destroys the folded structure of proteins. For a review of elementary chemistry, see eAppendix 1.
All cells share common chemical components:
Water, the fundamental solvent of life
Essential ions, such as potassium, magnesium, and chloride ions
Small organic molecules, such as lipids and sugars, that are incorporated into cell structures and that provide nutrition by catabolism
Macromolecules, such as nucleic acids and proteins, that contain information, catalyze reactions, and mediate transport, among many other functions
The details of bacterial chemistry emerged in the 1950s through pioneering studies by Fred Neidhardt at the University of Michigan and many colleagues. Cell composition varies with species, growth phase, and environmental conditions (as discussed in Chapters 4 and 5). Table 3.1 summarizes the chemical components of an E. coli cell during exponential growth.
TABLE 3.1
Molecules of a Bacterial Cell, Escherichia coli, during Balanced Exponential Growtha
Source: Modified from F. Neidhardt and H. E. Umbarger. 1996. Chemical composition of Escherichia coli, p. 14. In F. C. Neidhardt (ed.), Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed. ASM Press, Washington, DC.
Small molecules and ions. The E. coli cell consists of about 70% water, the essential solvent required to carry out fundamental metabolic reactions and to stabilize proteins. The water solution contains inorganic ions, predominantly potassium, magnesium, and phosphate. Inorganic ions store energy in the form of transmembrane gradients, and they serve essential roles in enzymes. For example, a magnesium ion is required at the active site of RNA polymerase to help catalyze the linking of ribonucleotides into RNA.
The cell also contains many kinds of small, charged organic molecules, such as phospholipids and enzyme cofactors. A major class of organic cations is the polyamines, molecules with multiple amine groups that are positively charged when the pH is near neutral. Polyamines balance the negative charges of the cell’s DNA and stabilize ribosomes during translation.
Macromolecules. Many cells have similar content of water and small molecules, but their specific character is defined by their macromolecules, especially their nucleic acids (DNA and RNA) and their proteins. DNA and RNA molecules can be isolated by size using agarose-gel electrophoresis, in which the negatively charged molecules migrate in an electrical field (see eAppendix 3). The total DNA plus RNA content of bacteria is relatively high, nearly 8% for E. coli—much higher than in multicellular eukaryotes. For microbes, the high nucleic acid content allows the cell to maximize reproduction of its chromosome while minimizing cell resources.
A high proportion of nucleic acids in food is a problem for us, because humans lack the enzymes to digest the uric acid waste product of nucleotides. That’s why we cannot eat most kinds of bacteria as a major part of our diet. Nevertheless, we do consume bacteria within vegetables, because all plants have bacteria growing within their transport tissues.
The cell’s genomic DNA directs expression of its proteins (discussed in Chapters 7–10). A given cell uses different genes to make different proteins, depending on environmental conditions such as temperature, nutrient levels, and entry into a host organism. Individual proteins are made in very different amounts, from 10 per cell to 10,000 per cell. The proteins expressed by a cell under given conditions are known collectively as a proteome. Other kinds of macromolecules are found in the cell wall and outer membrane. The bacterial cell wall consists of peptidoglycan, an organic polymer of peptide-linked sugars that constitutes nearly 1% of the cell mass, approximately the same mass as that of DNA. Peptidoglycan limits the volume of the enclosed cell, so water rushing in generates turgor pressure. This investment of biomass in the cell wall shows the importance (for most species) of maintaining turgor pressure in dilute environments, where water would otherwise enter by osmosis, causing osmotic shock (see eAppendix 2 and Section 3.2).
Thought Question
3.1 Which chemicals do we find in the greatest number in a bacterial cell? The smallest number? Why does a cell contain 100 times as many lipid molecules as strands of RNA?
ANSWER ANSWER
The chemicals that occur in the greatest number in a prokaryotic cell are inorganic ions (250 million/cell). They are also the smallest in size. DNA molecules are found in the lowest number (one large molecule, branched during replication). A prokaryotic cell contains 100 times as many lipid molecules as strands of RNA because lipids are small structural molecules, highly packed. They are a major component of the cellular membrane. RNA molecules are long macromolecules that either are packed into complexes (such as ribosomal RNA) or are temporary information carriers (messenger RNA), present only as needed to make proteins.
Cell Fractionation
Cell fractionation (Fig. 3.3) is how we separate cell components such as membranes, ribosomes, and flagella. We can study these isolated parts in detail, though we lose information about interactions with other parts of the cell. Cell fractionation also provides purified proteins that act as antigens for candidate vaccines. For example, a vaccine against Neisseria meningitidis type B (meningococcus) contains a highly immunogenic outer membrane protein.
More information
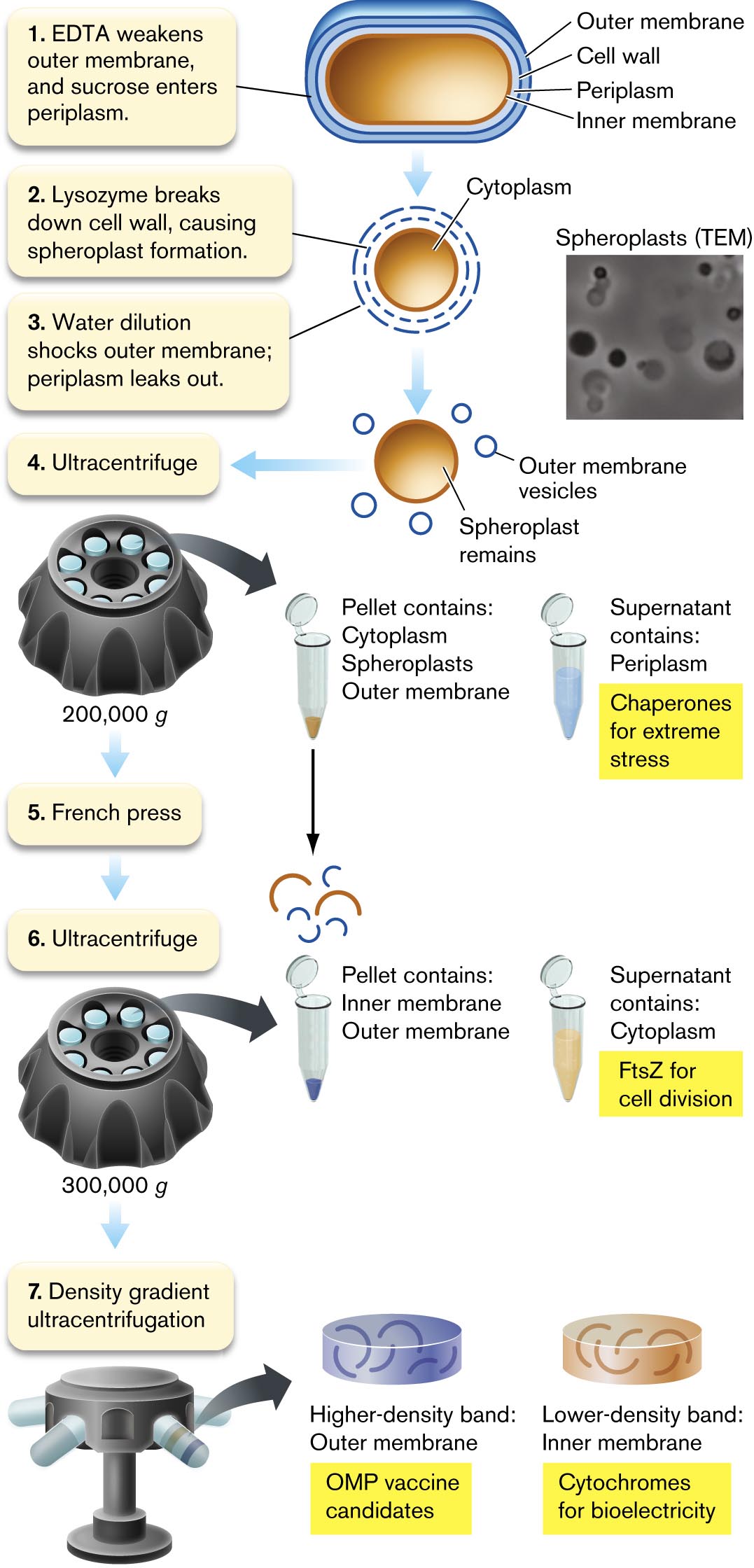
An illustration describes the process of fractionation of Gram-negative cells. The fractionation has 7 steps. 1. E D T A weakens outer Membrane, and sucrose enters periplasm. The cell consists of an outer membrane, cell wall, periplasm, and inner Membrane. 2. Lysozyme breaks down the cell wall, causing spheroplast formation. The center region of the cell is labeled as the cytoplasm. The cell wall is shown broken up. 3. Water dilution shocks the outer Membrane; periplasm leaks out. An image of the sphere shown is labeled as Spheroplast remains inside of the cell, and small circles scattered around the sphere is labeled as outer membrane vesicles. An inset shows a microscopic view of spheroplasts with several dark circles. 4. Ultracentrifuge. A narrow tube is shown. Ultracentrifugation at 200,000 g force. The pellet contains Cytoplasm, Spheroplasts, and outer Membrane. The supernatant contains periplasm and chaperones for extreme stress. 5. French press. 6. Ultracentrifuge. Ultracentrifugation at 300,000 g force. The pellet contains the inner Membrane and outer Membrane. The supernatant contains cytoplasm and F t s Z for cell division studies. 7. Density gradient ultracentrifugation. The higher-density band contains outer Membrane and O M P vaccine candidates. The lower-density band contains an inner membrane and cytochromes for bioelectricity.
FIGURE 3.3 ■Fractionation of Gram-negative cells. Cell periplasm fills with sucrose, and lysozyme breaks down the cell wall. Dilution in water causes osmotic shock to the outer membrane, and periplasmic proteins leak out. Subsequent centrifugation steps separate the proteins of the periplasm, cytoplasm, and inner and outer membranes. EDTA = ethylenediaminetetraacetic acid; OMP = outer membrane protein.LARS D. RENNER AND DOUGLAS B. WEIBEL. PNAS108:6264
How can we disassemble a cell to isolate its parts? Early-twentieth-century microbiologists wondered how to separate cell parts without all the molecules mixing together. As we learned in Chapter 1, the answer was ultracentrifugation. Ultracentrifugation (that is, rotation at high speed) subjects a suspension of molecules to high g force by centripetal acceleration. The g force separates molecules by weight and density, as explained in eAppendix 3. Mary Jane Osborn (1927–2019) at the University of Connecticut Health Center discovered that the inner and outer membranes of Gram-negative bacteria have different densities, and thus can be separated by density gradient ultracentrifugation.
Cell fractionation requires techniques that lyse (break open) the cell. The lysis method must generate enough force to separate the membrane lipids (held together by hydrophobic force) but not enough to disintegrate complexes of protein and RNA. For a Gram-negative cell, the method requires further specificity to separate “compartments” containing different sets of proteins: the inner and outer membranes, and the aqueous cytoplasm and periplasm.
Cell wall lysis and spheroplast formation. First we permeabilize the bacterial outer membrane by removal of Mg2+ and Ca2+ ions (Fig. 3.3, step 1). The permeabilized membrane now allows sucrose to cross. Sucrose fills the periplasm, maintaining an osmotically stable solution.
Next, lysozyme cleaves peptidoglycan and thus breaks down the cell wall (step 2). Lacking the turgid cell wall, the cell swells into a sphere called a spheroplast. Spheroplasts can be seen by transmission electron microscopy (TEM).
To isolate the periplasmic contents, the spheroplasts are transferred to distilled water (Fig. 3.3, step 3). Water rushes in through the EDTA-weakened outer membrane, causing osmotic shock of the periplasmic compartment, while the inner membrane remains intact. The periplasm leaks out into the extracellular medium, where its proteins can be collected. The periplasm is a source of interesting proteins such as sugar transporters and chaperones (proteins that help other proteins fold under stress).
Note that for different kinds of cells, other means of cell disruption may work better. Mild-detergent lysis can dissolve membranes without denaturing proteins. Alternatively, sonication is a way to lyse a cell by intense ultrasonic vibrations that are above the range of human hearing. For especially tough cells, such as cyanobacteria, a “bead beater” with microscopic glass beads can tear the cells open.
Ultracentrifugation. After osmotic shock, the spheroplasts undergo ultracentrifugation (Fig. 3.3, step 4) to separate the periplasmic contents from the other three types of cell compartments (cytoplasm, inner membrane, and outer membrane fragments, which form vesicles). The ultracentrifuge is a device in which tubes containing solutions of cell components are spun at very high speed. The high rotation rate generates centrifugal forces strong enough to separate subcellular particles (see eAppendix 3). The particles are collected in fractions of sample from the tube, and the fractions are observed by electron microscopy.
Spheroplast lysis. The pellet can now be further processed by a French press (Fig. 3.3, step 5), a device that squeezes cell contents through a narrow tube to break open the membranes. The broken membranes coalesce into tiny vesicles. A second step of ultracentrifugation now pellets the inner and outer membrane vesicles, while removing the cytoplasm in the supernatant (liquid above the pellet; step 6). The cytoplasm provides many types of proteins for study, such as FtsZ, as well as complexes (multiprotein structures) such as the ribosomes and DNA polymerases.
Finally, the membrane fraction contains a mixture of very different components of the inner membrane (such as electron transport proteins) and the outer membrane (such as cell-surface proteins that we could use to make vaccines). We can separate the inner and outer membranes by density gradient ultracentrifugation (Fig. 3.3, step 7). The gradient of solution density is generated by forming a gradient of sucrose concentration. In the gradient, the inner and outer membrane vesicles separate because of differences in their density, not their particle size. The lower-density fractions contain inner membrane vesicles, whereas the higher-density fractions contain outer membrane vesicles.
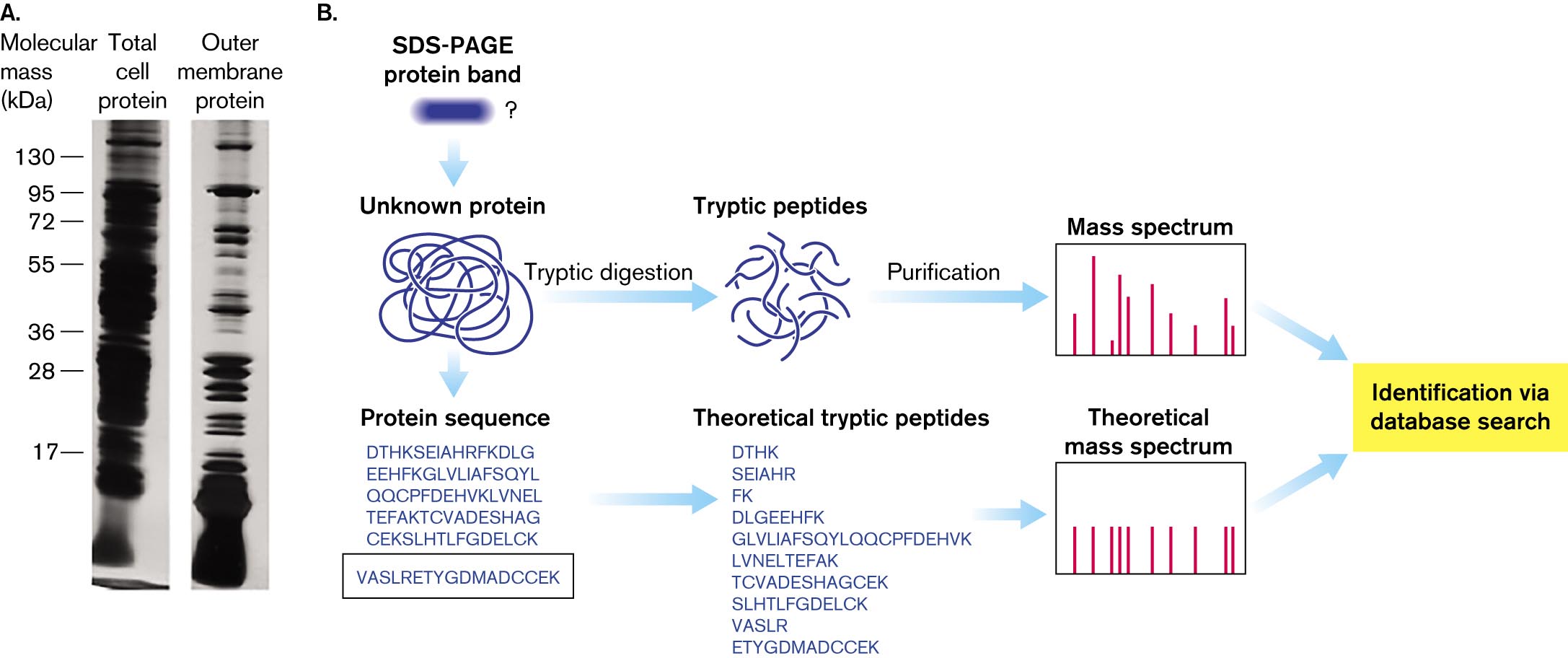
The membrane vesicle proteins are analyzed on electrophoretic gels (Fig. 3.4). We can identify the protein bands on the gel by enzyme digestion and mass spectrometry. The enzyme trypsin cleaves proteins only at aminoacyl residues lysine or arginine. The position of lysine or arginine residues is defined by the overall protein sequence; thus, the cleavage patterns generate peptides of defined composition.
More information
A photo of two protein gel electrophoresis bands and a diagram of protein mass spectrum analysis. The photo of gel electrophoresis bands comparing total cell proteins to outer membrane proteins. There are markers at several molecular mass units, in kilo Daltons. The total cell protein gel is completely dark. Individual bands are indistinguishable. The outer membrane protein gel contains several distinguishable bands. The diagram of protein mass spectrum analysis begins from a S D S-P A G E protein band analysis that leads an unknown protein. Through a several step process, the protein can be identified.
A photo of gel electrophoresis bands comparing total cell proteins to outer membrane proteins. There are markers at molecular mass units, in kilo Daltons, of 17, 28, 36, 55, 72, 95, and 130. The total cell protein gel is completely dark. Individual bands are indistinguishable. The outer membrane protein gel contains several distinguishable bands.
A diagram of protein mass spectrum analysis. It begins from S D S-P A G E protein band analysis that leads an unknown protein. The unknown protein is denatured. The unknown protein can undergo tryptic digestion or the protein sequence can be identified. If the protein undergoes tryptic digestion, tryptic peptides are isolated. The peptides are then purified and analyzed with mass spectrometry. The protein can be identified via a database search following mass spectrum analysis. If the protein sequence is identified, theoretical tryptic peptides can be listed. These theoretical peptides can be analyzed with mass spectrometry. The protein can then be identified via a database search following mass spectrum analysis.
FIGURE 3.4 ■Protein analysis.A. Gel electrophoresis of total cell proteins compared to outer membrane proteins from cell fractionation. B. Outer membrane proteins are identified by tryptic digestion and mass spectrum analysis. The resulting peptide sequence is compared with those predicted from genomic data. SDS-PAGE = SDS polyacrylamide gel electrophoresis.M. THEIN ET AL. 2010. J. PROTEOME RES.9:6135
The mass of each defined peptide is then determined by mass spectrum analysis (mass spectrometry). Mass sizes are used to predict the peptide amino-acid sequence composition. The predictions are compared with a database of protein sequences that the organism’s genome predicts. Thus, a protein’s sequence data can be used to discover the gene that encodes it.
Thought Question
3.2 Suppose we wish to isolate multidrug efflux pumps, which are protein complexes that span the envelope from inner membrane to outer membrane. How might we modify the cell fractionation procedure to achieve such isolation?
ANSWER ANSWER
A multidrug efflux complex such as AcrAB-TolC crosses the entire envelope, in order to completely expel antibiotics. The complex has subunits that span both inner and outer membranes, complicating the cell fractionation. It is possible that the complex might associate more strongly with one membrane or the other, so it could be found primarily in one of the membrane fractions. A fluorescent antibody could identify fractions that contain the complex. Mild-detergent treatment of the fractions could strip away the membrane. Alternatively, the whole-membrane prep could be treated with mild detergent. The protein fraction could then be centrifuged through a sucrose gradient; because the subunits of the complex have very specific size and density, they would be concentrated in distinct fractions. Electron microscopy could confirm the fraction containing the subunits, which could be reconstituted as a complex.
A limitation of cell fractionation is that it provides little information about processes that require an intact cell, such as cell division. How can we remove or alter a part of a cell without breaking it open? An approach that is complementary to cell fractionation is genetic analysis. In genetic analysis, we can mutate a strain so as to lose or alter a gene; then we select mutant strains for loss of a given function. The phenotype of the mutant cell may yield clues about the function of the altered part, such as the bacterial cytoskeleton (see Section 3.3). Genetics and genetic analysis are discussed further in Chapters 7–12.
To Summarize
Bacterial cells are protected by a thick cell envelope. The envelope includes a cell membrane (plasma membrane) and a peptidoglycan cell wall.
A Gram-negative cell includes an outer membrane. The cell membrane of a Gram-negative cell is called the inner membrane. Between the outer membrane and inner membrane lies the periplasmic space, containing the cell wall.
Bacteria are composed of nucleic acids, proteins, phospholipids, and other organic and inorganic chemicals. Proteins in the cell vary, depending on the species and environmental conditions.
The bacterial cytoplasm is highly structured. DNA replication, RNA transcription, and protein synthesis occur coordinately within the cytoplasm.
Cell fractionation isolates cell parts for structural and biochemical analysis. The compartments of a Gram-negative cell can be separated by cell lysis and ultracentrifugation.
Cell compartments contain different types of proteins. The outer membrane and inner membrane, as well as the cytoplasm and periplasm, each contain distinct types of protein not found in the other compartments. Outer membrane proteins may be good candidate antigens for a vaccine.
Microscopy reveals cell structure. Transmission electron microscopy and cryo-electron microscopy show how cell parts fit within the cell as a whole.
A structure external to the cell membrane, such as the cell wall or outer membrane of a bacterium. For a virus, the envelope is a membrane enclosing the capsid or core particle.
A filamentous structure for motility. In prokaryotes, a helical protein filament attached to a rotary motor; in eukaryotes, an undulating membrane-enclosed complex of microtubules and ATP-driven motor proteins.
A filamentous structure for motility. In prokaryotes, a helical protein filament attached to a rotary motor; in eukaryotes, an undulating membrane-enclosed complex of microtubules and ATP-driven motor proteins.
Two layers of phospholipids; the hydrocarbon fatty acid tails face the interior of the bilayer, and the charged phosphate groups face the cytoplasm and extracellular environment. The cell membrane is a phospholipid bilayer.
A machine that subjects samples to high centrifugal forces and can be used to separate subcellular components.
Endnotes
Values shown are for a hypothetical “average” cell cultured with aeration in glucose medium with minimal salts at 37°C. Return to reference a
The total weight of the cell (including water) is about 10⁻12 gram (g), or 1 picogram (pg). Return to reference b
The number of different kinds is difficult to estimate for proteins and mRNA because some genes are transcribed at extremely low levels and because proteins and RNA include kinds that are rapidly degraded. Return to reference c
In rapidly growing cells, cell fission typically lags approximately one generation behind DNA replication; hence, two identical DNA copies per cell. Return to reference d
In rapidly growing cells, cell fission typically lags approximately one generation behind DNA replication; hence, two identical DNA copies per cell. Return to reference d

 ANSWER
ANSWER ANSWER
ANSWER