Describe the distinctive traits of ciliates and apicomplexan parasites.
Outline the complex life cycle of Plasmodium parasites that cause malaria.
Describe the diseases caused by hemoflagellates, giardia, and trichomonads.
Amebas
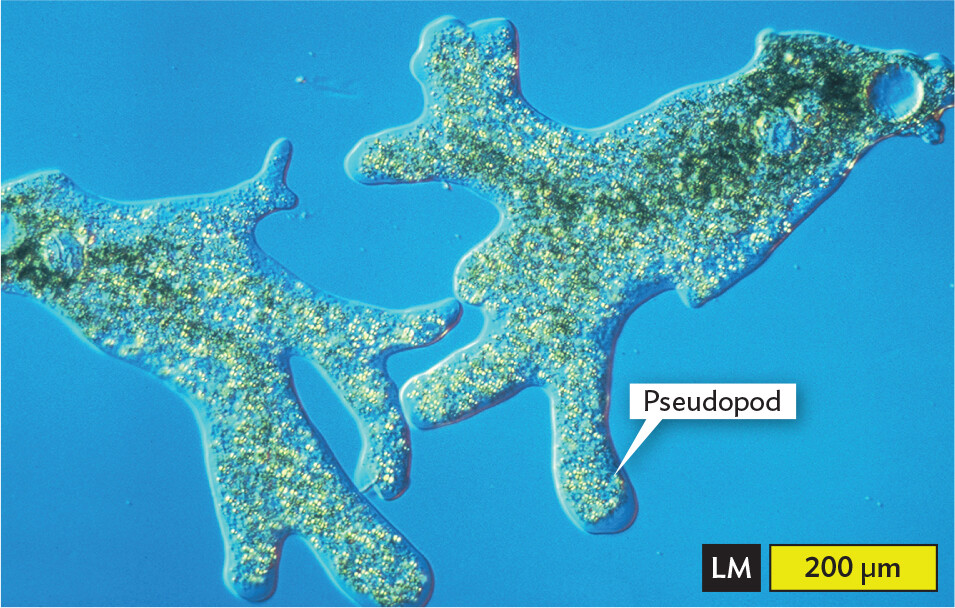
Amebas (also spelled “amoebas”) are voracious predators, consuming other protists and bacteria (Figure 11.9). Some kinds of amebas can reach several millimeters in size—these single-celled life forms are large enough to eat small invertebrates. Most amebas are free-living in soil or water, where they play major roles in the food web of soil and aquatic ecosystems.
Figure 11.9 Amebas
More information
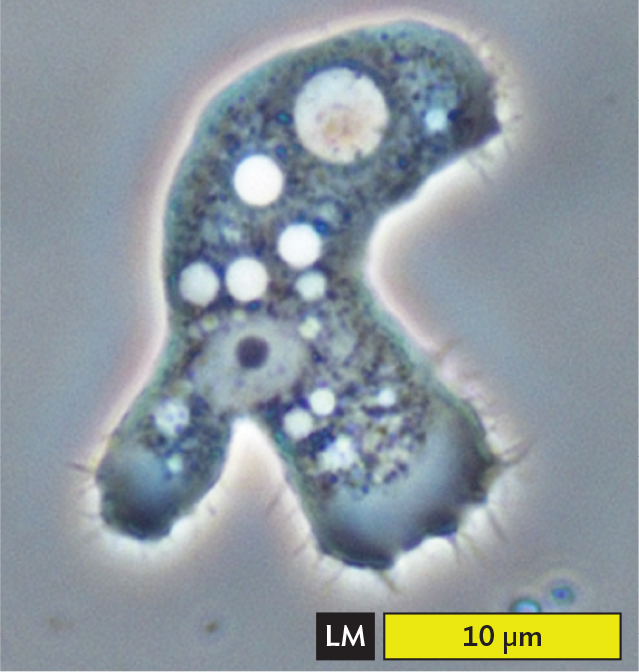
A light micrograph of Amoeba proteus. The micrograph shows a translucent irregular shaped cell that has formed branched lobe like structures. These are the pseudopods. Numerous dot like structures and sphere-like structures are present in the cytoplasm. The center body part of the amoeba appears green. There is one larger blue sphere at one end of the amoeba away from most pseudopods. The width of the pseudopod is approximately 60 micrometers. Thicker portions of the cell body have a width of about 200 micrometers.
A.Amoeba proteus extends its lobed pseudopods.More information
A light micrograph of the structure of a foraminiferan. The micrograph shows what appears as three flattened sphere like structures arranged one over the other, forming a single organism’s body. Long, thin needle like structures extend all around the organism’s shell like body. They are pseudopods. The size of the foraminiferan’s body is approximately 450 micrometers. Its pseudopods extend around 500 micrometers or more out from the body.
B. A foram (foraminiferan) extends needlelike pseudopods through its shell of calcium carbonate.
There are two main groups of amebas: the lobed amebas, with large, bulky pseudopods (Figure 11.9A); and the filamentous amebas, with thin, needlelike pseudopods (Figure 11.9B). Some amebas with needlelike pseudopods, such as the foraminifera (singular, foraminiferan; also called forams), form inorganic shells made of calcium carbonate (limestone). Fossil shells of foraminifera are common in rock formations derived from ancient seas. Foraminifera formed the white cliffs of Dover in Britain and the stone used to build the Egyptian pyramids.
The lobed amebas (the more familiar kind) have pseudopods that extend lobes of cytoplasm through cytoplasmic streaming. Cytoplasmic streaming is the flow of nutrients and other components through the cytoplasm, mediated by actin proteins. Some amebas are slime molds, in which individual amebas converge to form a fruiting body. Other ameba species, however, cause diseases such as dysentery and meningitis.
CASE HISTORY 11.2
Spring Break Surprise
More information
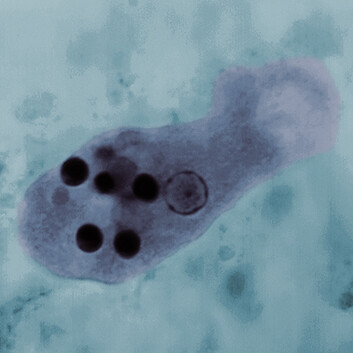
A light micrograph of Entamoeba histolytica. The micrograph shows an oval shaped sac like structure with numerous dark circles within. These circles are the ingested red blood cells. A larger round clear structure with a thin dark border is the nucleus.
Brianna, a 22-year-old college student, had returned recently from spring break at the beach in Acapulco, Mexico. She presented at the college clinic complaining of severe abdominal cramps and fever. She also had nausea and bloody diarrhea with mucus. The nurse practitioner collected stool samples and sent them to the laboratory for analysis. Cultures were negative for bacterial pathogens. A trichrome stain was performed, revealing amebas of the trophozoite (active feeding) stage (Figure 11.10A). The stained amebas contained ingested red blood cells (RBCs). The appearance of the amebas was characteristic of Entamoeba histolytica, the cause of amebic dysentery (bloody diarrhea), or intestinal amebiasis. Brianna was treated with metronidazole for 7 days, followed by paromomycin for 7 days, and her symptoms resolved.
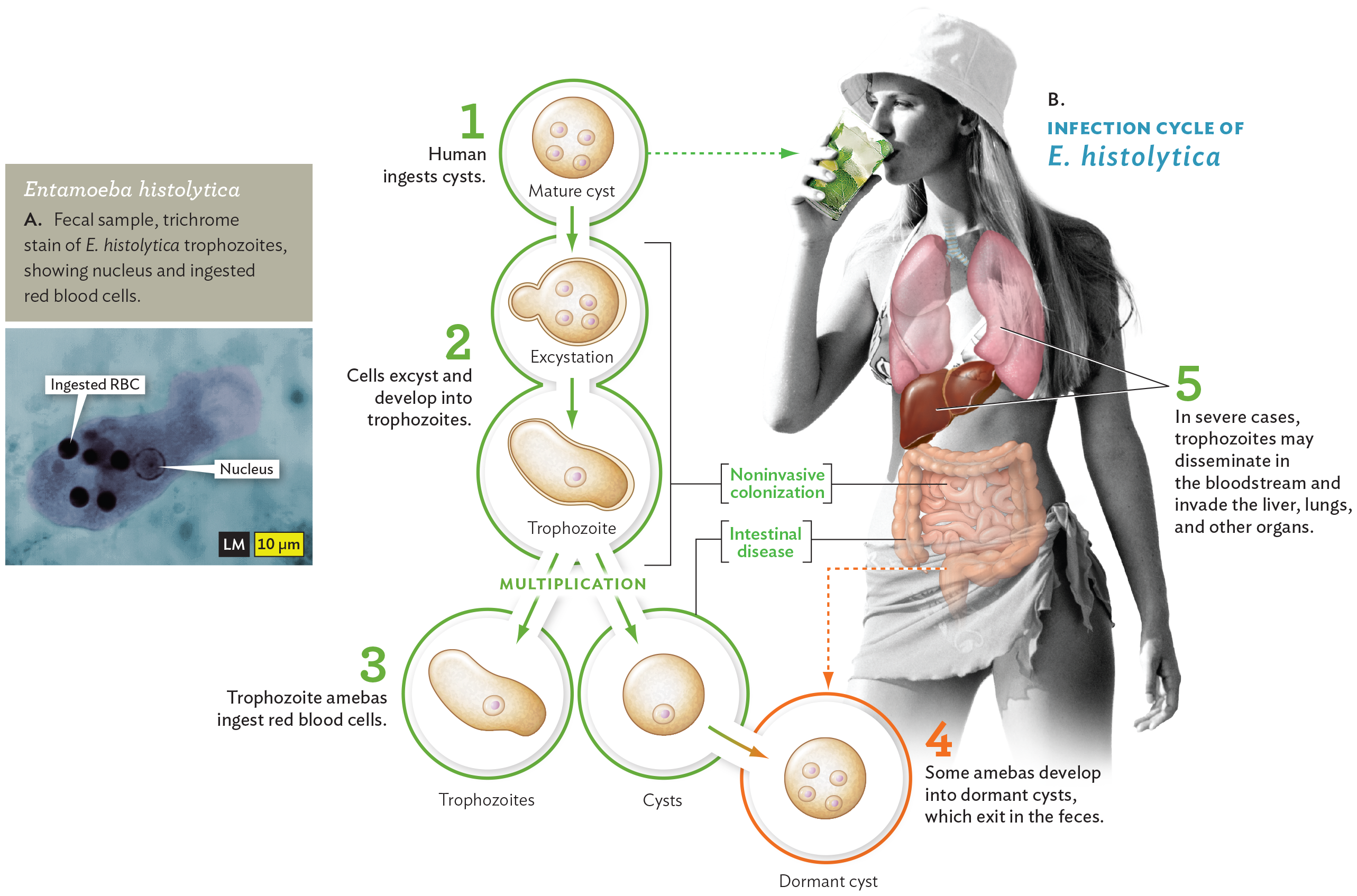
Figure 11.10Entamoeba histolytica
More information
A light micrograph of Entamoeba histolytica and a diagram of the infection cycle of Entamoeba histolytica. The light micrograph shows an irregularly shaped E histolytica trophozoite. The trophozoite contains a nucleus and several ingested red blood cells. The ingested red blood cells stain darkly. The diagram of the infection cycle contains 5 steps. There is an inset photo of a human consuming an iced drink with several illustrated organs overlayed. In the first step, the human ingests cysts through the drink. The mature cysts have four nuclei. In the second step, the cells excyst and develop into trophozoites. These steps are noninvasive colonization, which occurs in the small intestine. Excystation occurs via budding from the mature cyst. The trophozoite is irregularly shaped and has a single nucleus. In the third step, trophozoite amebas ingest red blood cells. The trophozoite multiplies into trophozoites and immature cysts. This is considered intestinal disease and occurs in the colon. In the fourth step, some amebas develop into dormant cysts which exit in the feces. This occurs in the sigmoid colon and rectum. In the fifth step, in severe cases, trophozoites may disseminate in the bloodstream and invade the liver, lungs and other organs.
Entamoeba histolytica is a serious cause of intestinal illness in much of the developing world, where it can lead to malnutrition and death. The amebas are usually ingested in the form of cysts, dormant cells encased in a tough coating (Figure 11.10B). Cysts can remain viable for many weeks in contaminated water supplies, such as those from which Brianna drank. The ingested cysts reach the intestine, where they “excyst” and develop into trophozoites, amebas with pseudopods that phagocytose prey. In the intestinal lumen and in the bloodstream, the amebas ingest red blood cells (RBCs). As the amebas run short of food, they develop into dormant cysts, which exit in the feces—a “fecal-oral” route of transmission. In severe cases, however, the trophozoites may invade the liver, lungs, and other organs.
The symptoms of dysentery can be caused by a number of different viruses and bacterial pathogens, which require different kinds of treatment; thus, the pathogen must be identified correctly. Common bacterial pathogens such as Shigella may be detected by immunological tests. But to diagnose the presence of amebas and other intestinal parasites requires microscopy because their cell size and shape are distinctive. A procedure called Wheatley’s trichrome stain reveals the parasites among human cells and fecal material (see Figure 11.10A). The trichrome stain involves treatment with iodine-alcohol solution, followed by a triple-stain solution. The cytoplasm of protozoa stains light purple or green, whereas the nuclear membrane and ingested RBCs stain deep purple. Cyst forms of the ameba stain bluish purple. Nevertheless, it must be noted that the closely related nonpathogen Entamoeba dispar is indistinguishable from E. histolytica under the microscope and is found ten times more often, so it is possible to miss the real pathogen.
Amebic dysentery is best avoided by careful choice of fluid consumption. When traveling in regions with limited sanitation, it is important to drink only filtered or bottled water. Avoid fresh produce that may be washed in contaminated water.
In most cases, an otherwise healthy host can eliminate amebic infection within 2 weeks. Repeated infections, however, may cause malnutrition. Sometimes the infection leads to chronic carrier status, and occasionally infection becomes systemic, involving the liver, lungs, and other organs. Systemic infection by amebas is difficult to treat and can be fatal. So, where good health care is available, cases of intestinal amebiasis are treated early with anti-amebic agents. The drug paromomycin specifically targets the small-subunit ribosomal RNA of microbial eukaryotes. An alternative drug, metronidazole, targets anaerobic protozoa and bacteria.
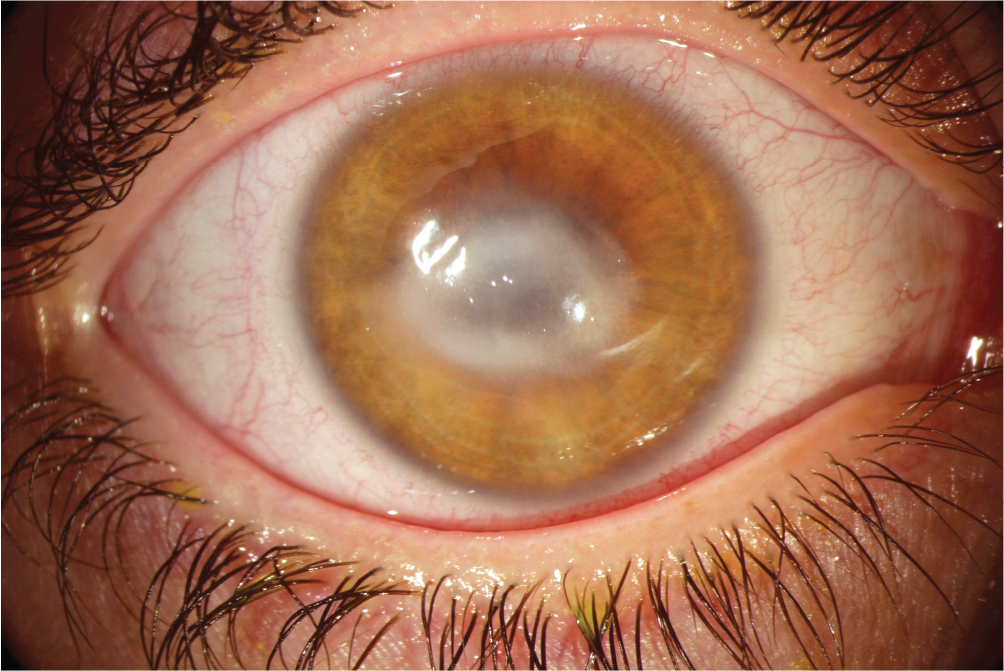
Other diseases can involve different species of amebas. For example, primary amebic meningoencephalitis, an infection of the brain (often fatal), can be caused by the amebas Naegleria fowleri and Balamuthia mandrillaris. The amebas may enter the body through the nose while a person is swimming in a contaminated pool. Free-living soil amebas such as Acanthamoeba may contaminate contact lens cleaning solutions, causing keratitis, an infection of the cornea (Figure 11.11). Wearers of reusable contact lenses have a keratitis infection rate of 2 per 1,000. Finally, free-living amebas may play host to bacterial pathogens such as Legionella pneumophila, the cause of legionellosis, an often-fatal form of pneumonia (discussed in Section 20.3). Amebas carrying intracellular L. pneumophila often contaminate water supplies, air ducts, and even dental irrigation solutions. The host amebas enable the pathogen’s persistence and transmission to human hosts.
Figure 11.11Amebic Keratitis
More information
A photo of an Acanthamoeba infection in the eye. The photo shows the Acanthamoeba infected keratin layers of the eye. Thick white clouded layers cover the eye so that the entire pupil is obscured. The sclera, surrounding the iris, is red and irritated.
A.Acanthamoeba infects the keratin layers of the cornea, which becomes inflamed and clouded.More information
A light micrograph of Acanthamoeba. The micrograph shows a light blue irregularly shaped organism with short hair like structures surrounding it. Numerous vacuoles, which appear as small white circular structures, are present in the cytoplasm. The nucleus is a larger clear oval shaped structure with a dark central circular structure. There are large, clear blue blurry blobs at the edges of the organism. The length of the Acanthamoeba is approximately 16 micrometers.
B.Acanthamoeba.
Amebic keratitis commonly results from improperly cleaned contact lenses.
Predators and Parasites
What other protozoa have interesting ways to catch prey? Ciliates are covered with cilia (singular, cilium), short, hairlike organelles composed of microtubules (Figure 11.12). The beating cilia propel a stream of prey to the oral groove (mouth). Many ciliates are voracious predators that swallow prey nearly their own size (Table 11.3). Other predatory protozoa called flagellates possess pairs of flagella—long, whiplike organelles. Like cilia, protozoan flagella are composed of microtubules. Their whiplike motion is powered by ATP.
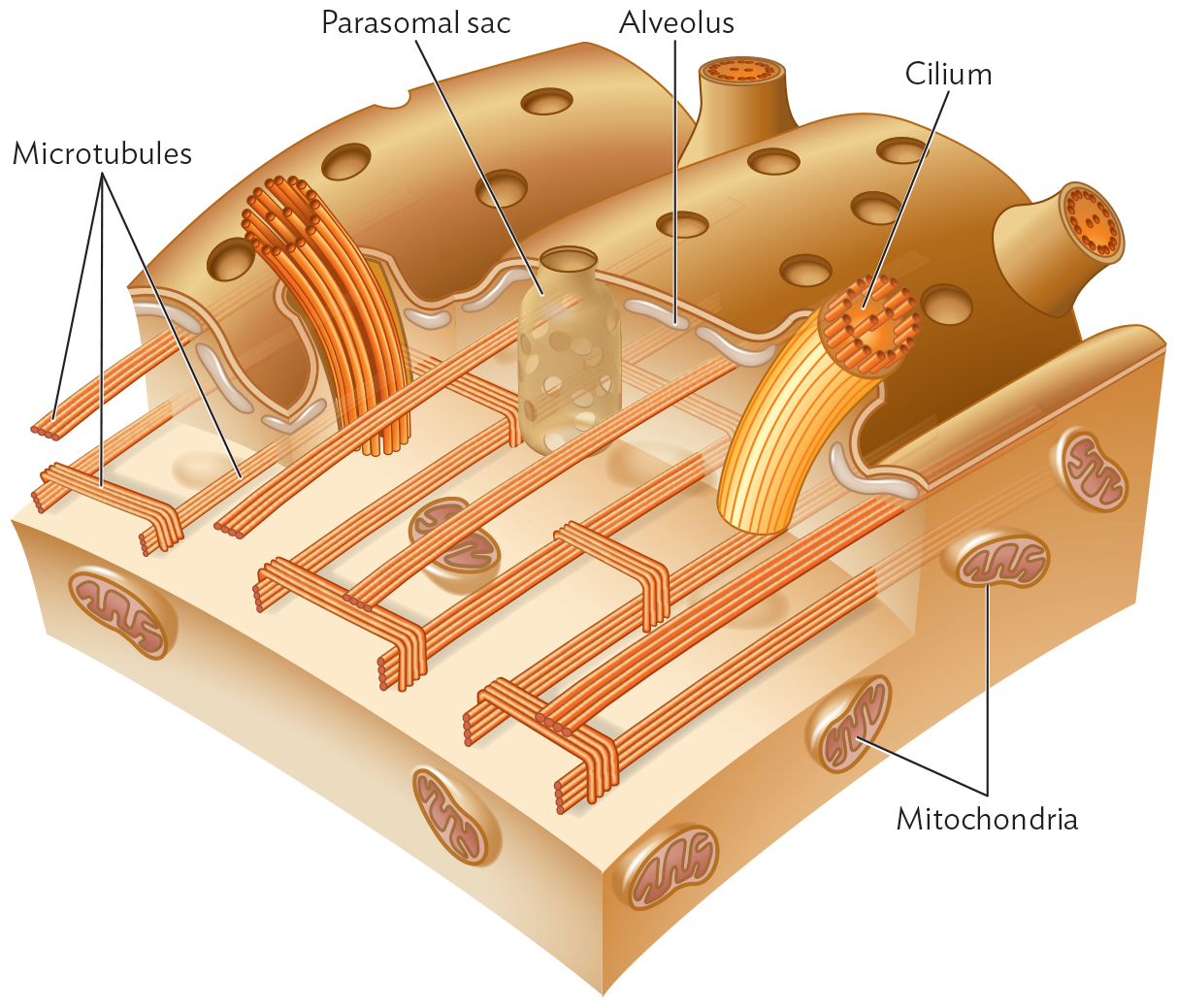
Figure 11.12 The Cortex of a Ciliate
More information
An illustration of the cortex of a ciliate. The illustration shows a flattened outer layer with small pores throughout. Below this layer, horizontal tubular sac like structures are the parasomal sacs. The tops of the sacs are the pores on the outer layer. There are also horizontal tubes just below the outer layer which are the alveoli. The third most layer at the bottom has strands made of bunches of tubular structures that show microtubules. These strands are placed horizontally across the length of the first layer. In some areas, these bunches of tubular structures cross perpendicularly over each other. Below this layer is a flattened three dimensional layer that contains oval shaped mitochondria. In two places there is a bundle of vertical tubular structures projecting up from the bottom and out of the top layer. These are the Cilia.
Beneath the pellicle (outermost cell layer) of a ciliated protist lie flattened sacs of fluid called alveoli. The cilia, each composed of microtubules, are rooted in a network of lateral microtubules. Parasomal sacs take up nutrients and form endocytic vesicles.
Table 11.3
Ciliates and Apicomplexans
Major Category and Genus
Habitat or Disease
Ciliates (ciliated protists)
Paramecium
Pond predator of algae and bacteria
Stentor
Stalked, ciliated predator; cilia draw prey into mouth; wastewater treatment community
Apicomplexans (parasites with apical complex; lack flagella or cilia; undergo complex life cycles)
Babesia
Babesiosis, blood infection spread by ticks
Cryptosporidium
Intestinal infection, opportunistic; waterborne
Plasmodium
Malaria; carried by mosquito vector
Toxoplasma
Toxoplasmosis; transmitted to humans by cats
Note: Eukaryotic flagella and cilia are very different from bacterial flagella, which are helical protein filaments. Bacterial helical flagella rotate, powered by the proton motive force across the cell membrane.
Ciliates
Ciliates are cilia-covered, heterotrophic protists that consume algae and smaller protists and are in turn consumed by amebas. A medically important ciliate is Balantidium coli, a cause of dysentery. Powered by ATP hydrolysis, the cilia beat in coordinated waves that maximize the efficiency of motility.
Cilia serve two functions:
Cell propulsion. Coordinated waves of beating cilia, usually covering the cell surface, propel the cell forward.
Food acquisition. By generating water currents into the mouth of the cell, a ring of cilia around the mouth brings food into the cell.
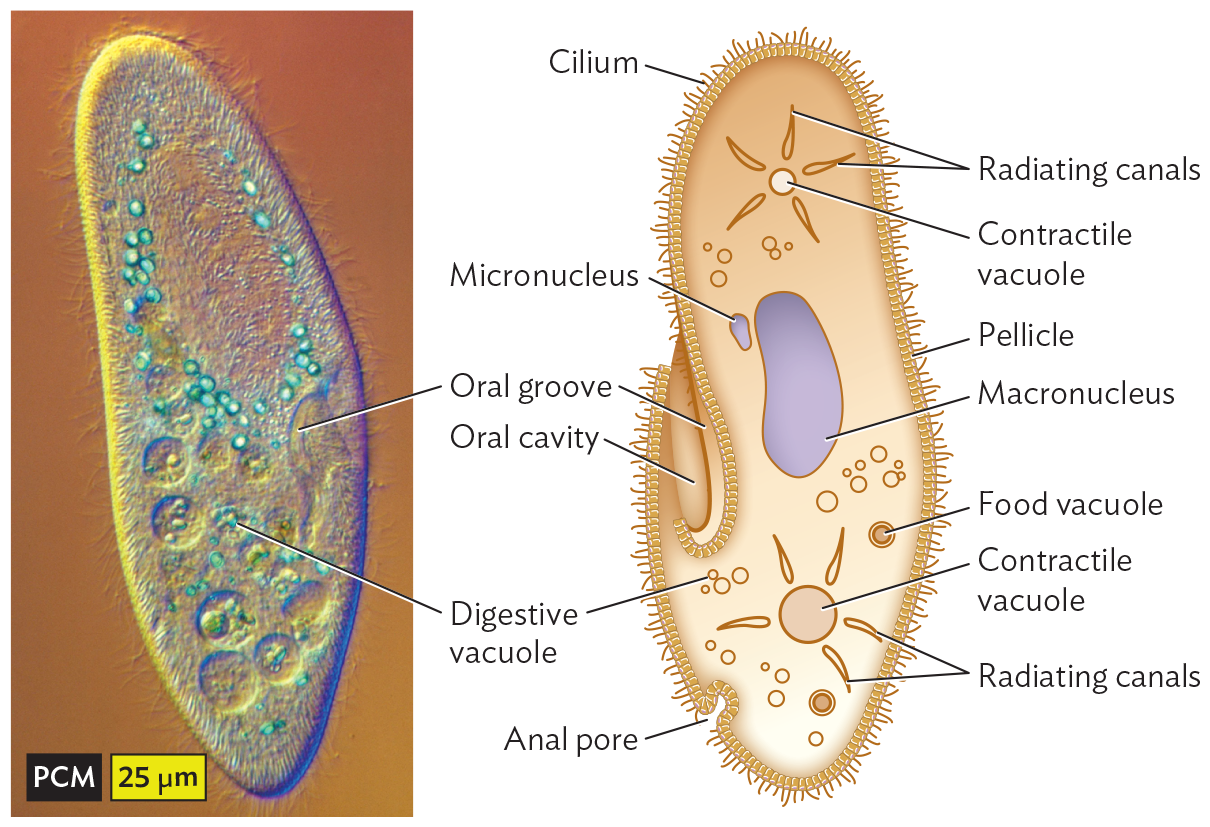
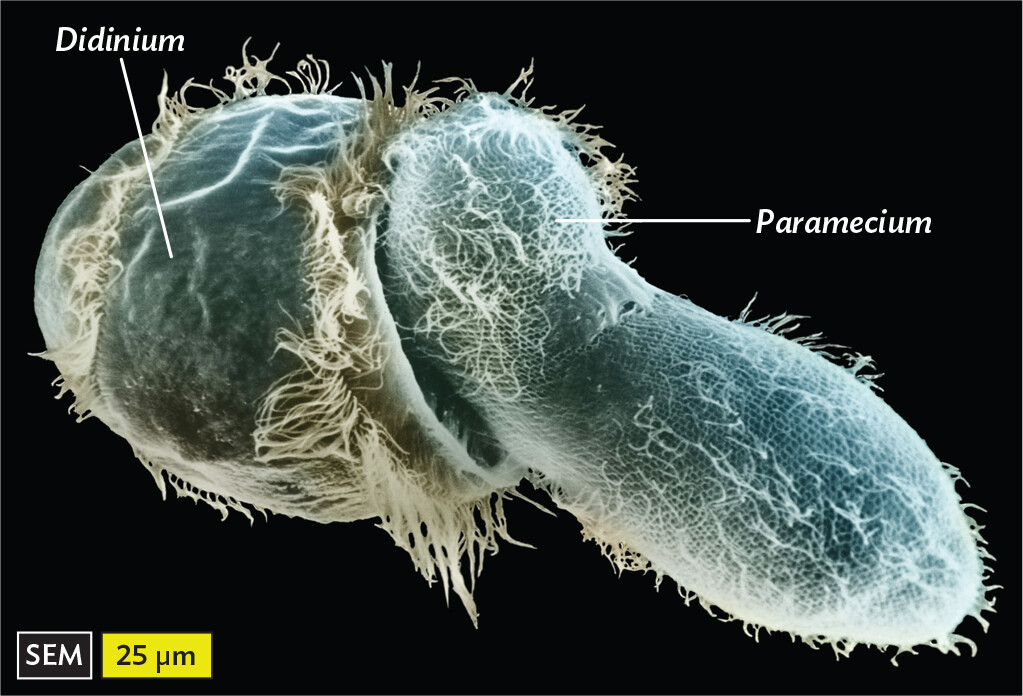
One of the best-studied ciliates is the paramecium (Figure 11.13A). Paramecia feed on bacteria by trapping them in the oral groove and then consuming them through enzymes within digestive vacuoles. Paramecia can also take up smaller particles through endocytosis by specialized pores in their cortex called parasomal sacs. Paramecia in turn are consumed by larger ciliates, such as Didinium (Figure 11.13B).
Figure 11.13 Ciliate Predators
More information
A phase contrast micrograph of a paramecium. The micrograph shows a vertically elongated oval shaped structure. It contains small light blue spherical structures labeled as digestive vacuoles and an irregularly shaped depression on the right side of the cell is labeled as the Oral groove. The body of the paramecium contains many circular vacuoles and other organelles. Its exterior is covered by hair like projections. The length and the width at the center are approximately 200 micrometers and 50 micrometers, respectively.
A. A paramecium has digestive vacuoles and an oral groove to ingest prey.More information
A scanning electron micrograph of the ciliate Didinium consuming a Paramecium. The micrograph shows the 3 D oval structure of Didinium covered with fur like cilia. Its surface has wrinkles almost like paper and one of its sides is flat and concave. On this side, it is consuming the elongated oval shaped structure of Paramecium. The paramecium is covered with numerous cilia that appear like threads on a gridded surface. The length and the width at the center of paramecium are approximately 150 micrometers and 50 micrometers, respectively. The length of the Didinium is approximately 100 micrometers.
B.Didinium consuming Paramecium. Both are ciliates.
Ciliates have a complex cell form called “alveolate,” referring to the flattened vacuoles called alveoli (singular, alveolus) within the outer cortex (Figure 11.12). Some alveoli contain plates of stiff material, such as protein, polysaccharide, or mineral. Most alveolate protists possess other kinds of cortical organelles in addition to alveoli. These include extrusomes (organelles that extrude defensive enzymes or toxins), bands of microtubules for reinforcement, and whiplike cilia or flagella.
Without a rigid cell wall, how do ciliates maintain osmotic balance? Ciliates generally grow in aqueous solutions with salt and organic concentrations that are lower than those of their cytoplasm. Thus, water concentration outside is higher, and water tends to run into the cell. The excess water is expelled by a contractile vacuole, which withdraws water from the cytoplasm and contracts to expel it. Contractile vacuoles are widespread among protists and algae, but their mode of action has been studied mostly in paramecia. As water enters the cell from a low-solute environment, the contractile vacuole takes up the water through an elaborate network of intracellular channels and then contracts to expel the water through a pore, thus preventing osmotic shock and lysis (see Section 4.5).
Ciliates, like other eukaryotes, have nuclei containing chromosomes that undergo division by mitosis and generate gametes by meiosis. In a setup unique to ciliates, however, the cell maintains two types of nuclei: the micronucleus, whose genes maintain genetic fidelity for sexual reproduction; and the macronucleus, containing multiple gene copies that express gene products throughout the large cell. During asexual reproduction, both micronuclear and macronuclear chromosomes undergo replication.
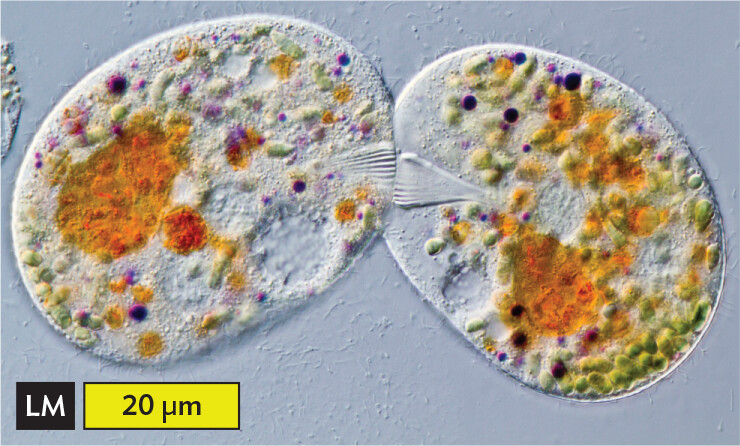
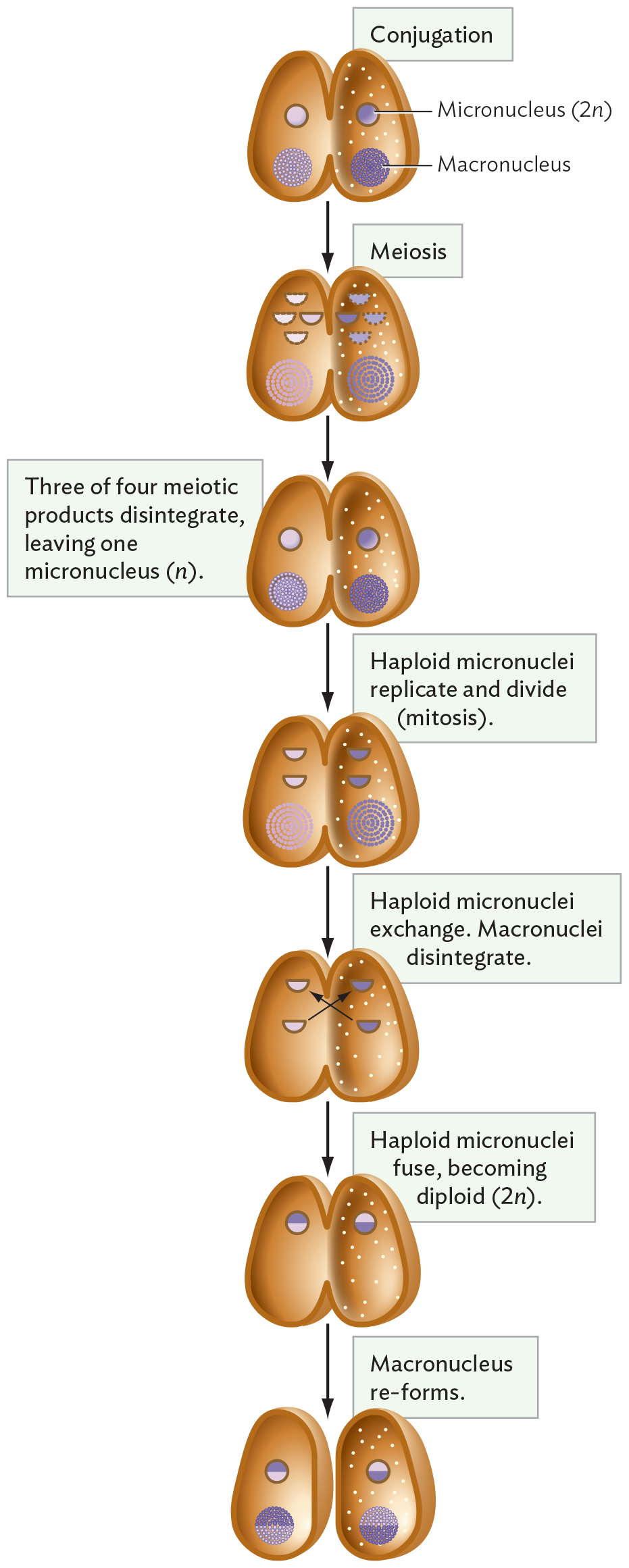
For sexual reproduction, a ciliate undergoes conjugation (Figure 11.14). During conjugation, two paramecia of opposite mating types form a cytoplasmic bridge. Within each conjugating cell, the 2n micronucleus undergoes meiosis to form four n gametic nuclei. (Meiosis was described in Section 11.1.) Three out of four of the gametic nuclei disintegrate, as does the macronucleus. The haploid micronuclei undergo mitosis, forming two daughter micronuclei. Daughter micronuclei from each cell are exchanged across the cytoplasmic bridge and then fuse with their respective counterparts, restoring a 2n micronucleus to each cell. The bridged cells come apart, and each micronucleus generates a new macronucleus. The new macronucleus now expresses gene products.
Figure 11.14 Conjugation
More information
A light micrograph of the process of conjugation in a pair of Ciliates. The micrograph shows two oval Ciliates touching one another on one end. There is a ridged triangle formation in each ciliate where they are touching. The flat sides of these triangles come in contact in the middle of the point of attachment. The ciliates each have large brown ovals inside and many small circular structures inside, these are food vacuoles and secretory granules. The diameter of one of the ciliates is approximately 40 micrometers.
A. Pair of ciliates conjugating. Colored objects are food vacuoles and secretory granules.
More information
A diagram of the processes of conjugation, meiosis, and mitosis between a pair of paramecia. There are two oval shaped Paramecia, longer on their vertical sides. During the conjugation stage, the two Paramecia are attached side to side. Their membranes are seen melding together and a gap forms between both membranes such that the cytoplasms become continuous. The right Paramecium has dot like structures in the cytoplasm, while the other does not have any dotted structures. Each Paramecium contains a small sphere shaped structure called a micronucleus two n and each has a large spherical structure made up of tiny concentric dots called a macronucleus. During the meiosis stage, the micronucleus divides into four half circles. The circles that make up the macronuclei are more spread out. In the next stage, only one sphere shaped structure of the micronucleus is found. A caption reads, Three of four meiotic products disintegrate, leaving one micronucleus, n. The circles that make up the macronuclei appear closer together again. In the next stage, the spherical micronucleus is shown divided into two half circles. The circles that make up the macronuclei are more spread out again. A caption reads, Haploid micronuclei replicate and divide by mitosis. In the next step, one of the two half circle structures in each Paramecium are exchanged between each paramecium. The macronuclei have disappeared. A caption reads, Haploid micronuclei exchange. Macronuclei disintegrate. In the next step, the spherical micronuclei are reformed by the combination of one of the original and one of the foreign micronuclei. The bottom half is color coded to relate to the original paramecium, and the top half is color coded to indicate it relates to the partner paramecium. A caption reads, Haploid micronuclei fuse, becoming diploid, two n. In the next step, the large spherical macronuclei reform. The bottom half is color coded to relate to the original paramecium, and the top half is color coded to indicate it relates to the partner paramecium. A caption reads, Macronucleus re forms.
B. Two paramecia of opposite mating types form a cytoplasmic bridge. The 2n micronucleus of each cell undergoes meiosis. Each macronucleus, as well as three out of four meiotic products, disintegrates. The haploid micronuclei undergo mitosis, forming two daughter micronuclei. Daughter nuclei from each cell are exchanged across the cytoplasmic bridge and then fuse with their respective counterparts, restoring 2n micronuclei. The cells come apart, and each micronucleus generates a new macronucleus.
Note: Distinguish the conjugation of ciliates (with exchange of micronuclei) from the very different process of bacterial conjugation (one-way transfer of DNA from a donor bacterium to a recipient).
Apicomplexans
Apicomplexans form a major group of parasites of humans and other animals (Table 11.3). Their name derives from the apical complex, a specialized structure that facilitates entry of the parasite into a host cell. Like the ciliates, apicomplexans are alveolates possessing an elaborate cortex composed of alveoli, pores, and microtubules. But as parasites, apicomplexans have undergone extensive reductive evolution, losing their flagella and cilia.
Parasitic protists may have life cycles involving multiple hosts. For example, the malaria parasite (Plasmodium) alternates between the human host and the Anopheles mosquito. Generally, one type of animal constitutes the definitive host, the host where the parasite matures and may reproduce sexually; in the case of malaria, the definitive host is the mosquito. The alternate host (the human in this case) is called the intermediate host. The intermediate host supports asexual proliferation of the parasite and is required for transmission to the next insect host.
Malaria. The best-known apicomplexan parasites are Plasmodium falciparum and Plasmodium vivax, the main causes of malaria, the most significant parasitic disease of humans worldwide (Figure 11.15A). P. falciparum and P. vivax are carried by mosquitoes, which transmit the parasite to humans when the insect’s proboscis (mouthparts) penetrates the skin. The disease is endemic in areas inhabited by 40% of the world’s population; each year malaria infects hundreds of millions of people and kills hundreds of thousands of African children. As people travel, malaria has increasingly showed up in the United States as well.
Figure 11.15Malaria
More information
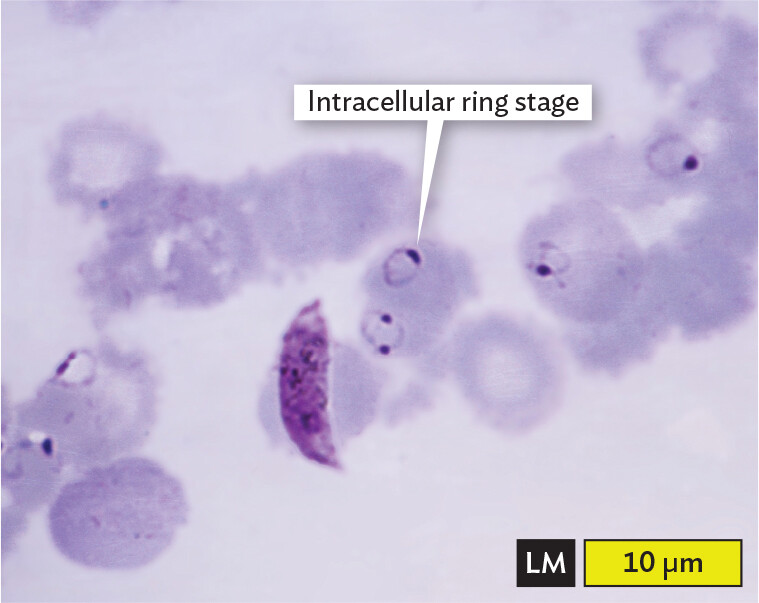
A light micrograph of Red Blood Cells infected by Plasmodium falciparum. The micrograph shows a blood smear. The red blood cells are disc shaped, with darker outer edges and pale staining centers. A few of the red blood cells contain ring like structures with a thick dot at one end. One of these rings is labeled intracellular ring stage. There is one long banana shaped cell that is stained the same shade as the thick dots on the rings. It lays across one of the red blood cells. It is almost 10 micrometers long. The size of the intracellular ring stage is about 2 micrometers across. Each red blood cell has a diameter of about 5 micrometers.
A. Red blood cells (RBCs) infected with Plasmodium falciparum, an apicomplexan parasite that causes malaria (stained purple with a dye that interacts with DNA).More information
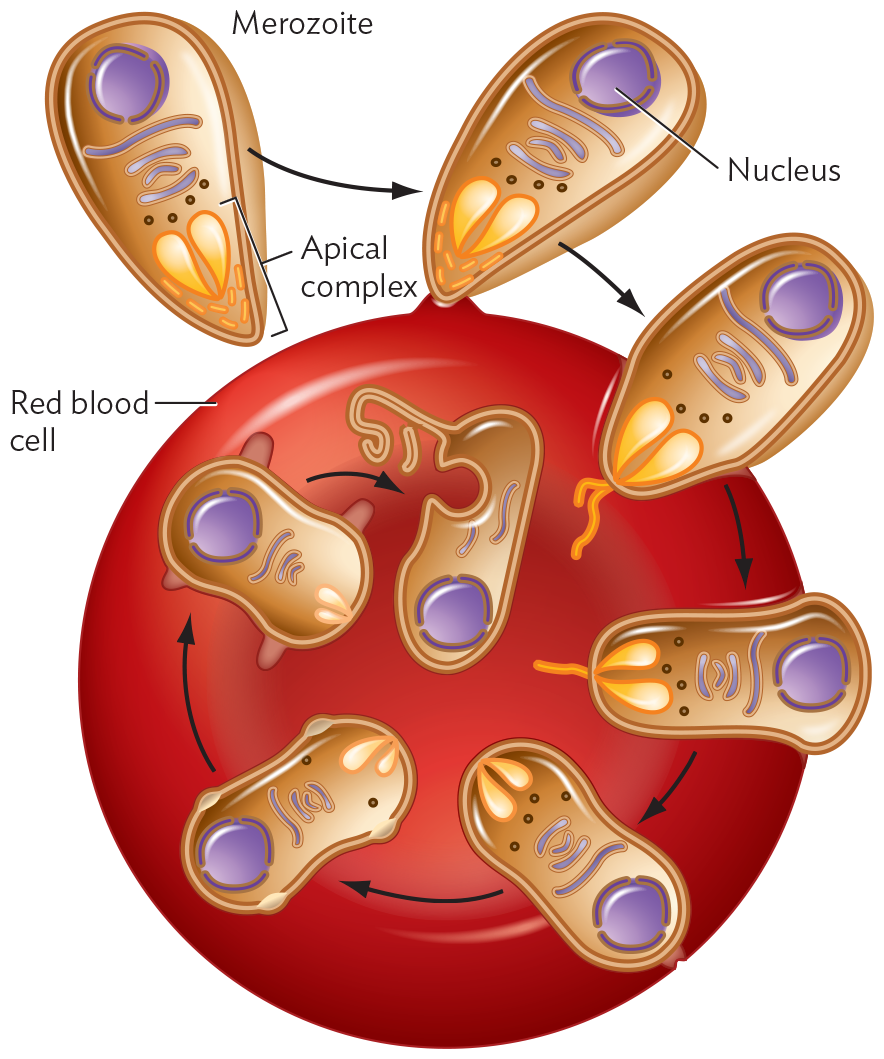
A diagram of the merozoite form of Plasmodium falciparum invading a Red Blood cell. The merozoite form of Plasmodium falciparum is tear drop shaped. The cell’s interior consists of a spherical nucleus, long tubules of the Golgi complex and small spherical ribosomes. The pointed end of the merozoite consists of an apical complex showing two teardrop shaped structures surrounded by tiny tubular structures. In the first stage, the merozoite attaches to the disc shaped Red Blood cell by its pointed end. On attachment, in the second stage, the apical complex contents are released outside the merozoite through the tubular structures. In the third stage, the contents start dissolving inside the red blood cell. The merozoite is now in a cylindrical shape. In the fourth stage, the merozoite is slipper shaped with a constricted middle region. There is no longer a tubular structure outside the cell. In the fifth and sixth stages, the merozoite membrane blebs outwards. The apical complex inside the merozoite shrinks. In the final stage, the merozoite is internalized into the R B C. There is no apical complex. The merozoite appears as an oval with a circular depression in the side and a long strand of membrane extending outwards.
B. The merozoite form of P. falciparum invades a red blood cell. The apical complex facilitates invasion and then dissolves as the merozoite transforms into an intracellular form.
The full life cycle of the malaria parasite includes developmental stages in the mosquito, followed by transmission to the human host, where the parasites undergo stages in the liver and in the blood. In the liver, the transmitted parasites develop into the “merozoite” form, which invades RBCs. The merozoite first contacts a red blood cell through interaction between its apical complex (the structure at the penetrating tip of the cell) and receptor proteins on the RBC (Figure 11.15B). The apical complex contains a pair of secretory organelles that are capped by a ring of microtubules and that inject enzymes that aid entry of the parasite. The tip of the apical complex penetrates the host cell, enabling secretion of lipids and enzymes that facilitate invasion. Eventually, the entire merozoite enters the host cell, leaving no traces of the parasite on the host cell surface. Thus, the internalized parasite (ring stage) becomes invisible to the immune system until its progeny burst out. Some progeny are then picked up by mosquitoes, where they undergo a sexual life cycle and are transmitted to new hosts.
The malarial parasites acquire resistance rapidly and no longer respond to drugs, such as quinine, that nearly eliminated the disease half a century ago. The life cycle and molecular properties of P. falciparum have been studied extensively for clues to the development of new antibiotics and vaccines. The life cycle of P. falciparum and the etiology of malaria are discussed in Chapter 21.
Toxoplasmosis. An apicomplexan parasite common in the United States is Toxoplasma gondii, a parasite carried by cats and transmissible to humans. Approximately 30% of the world’s population is infected with T. gondii. Fortunately, the infection is usually asymptomatic in humans because of our immune system, but the parasite can cross the placental barrier and harm a developing fetus. In rare cases, T. gondii can enter the brain, where it can alter host behavior. Some epidemiological evidence suggests a link between T. gondii infection and some cases of schizophrenia. The life cycle of T. gondii and the etiology of toxoplasmosis are discussed in Chapter 21.
Babesiosis. An emerging human pathogen in the United States is the blood parasite Babesia microti. B. microti is spread by deer ticks, the same agents that spread Lyme disease—yet another unfortunate risk of a walk in the northeastern woods. B. microti, however, may also be spread by blood contact and transfusions. As in malaria, the parasite multiplies within RBCs. The symptoms of B. microti infection can mimic those of Lyme disease (caused by the tick-borne bacterium Borrelia burgdorferi) and are often misdiagnosed. Babesiosis is discussed further in Chapter 21.
Waterborne apicomplexans. Several apicomplexans are emerging waterborne parasites. Cryptosporidium parvum infects the intestine of immunocompromised patients, causing watery diarrhea. An obligate intracellular pathogen, the organism generates a cyst form transmitted by the fecal-oral route. Cryptosporidium resists chlorination; it can cause major outbreaks of intestinal disease, such as the one in 2013 when nearly 3,000 people were sickened from municipal drinking water in Baker City, Oregon. Another cause of diarrhea is Cyclospora, also a danger for immunocompromised patients. Cyclospora causes occasional outbreaks from fecally contaminated produce.
Hemoflagellates
What other motile parasites cause human disease? Euglenids are a group of protists that have whiplike flagella and chloroplasts arising from secondary endosymbiosis, like the dinoflagellates do (see Section 11.4). Some euglenid taxa, however, lost their chloroplasts and evolved into obligate parasites called hemoflagellates (some of which are called trypanosomes). A hemoflagellate cell is elongated, with a single flagellum. Hemoflagellate cells have a unique organelle called the kinetoplast, which evolved as a specialized mitochondrion that provides energy for the flagellum.
Hemoflagellates cause devastating diseases in humans and animals, such as trypanosomiasis (Trypanosoma brucei; Figure 11.16A). Trypanosomiasis, also known as African sleeping sickness, is a major killer of humans and livestock; in some sub-Saharan countries, it is the second-biggest killer after AIDS. Other diseases caused by hemoflagellates include Chagas disease (Trypanosoma cruzi) and leishmaniasis (Leishmania donovani; see the chapter-opening case history).
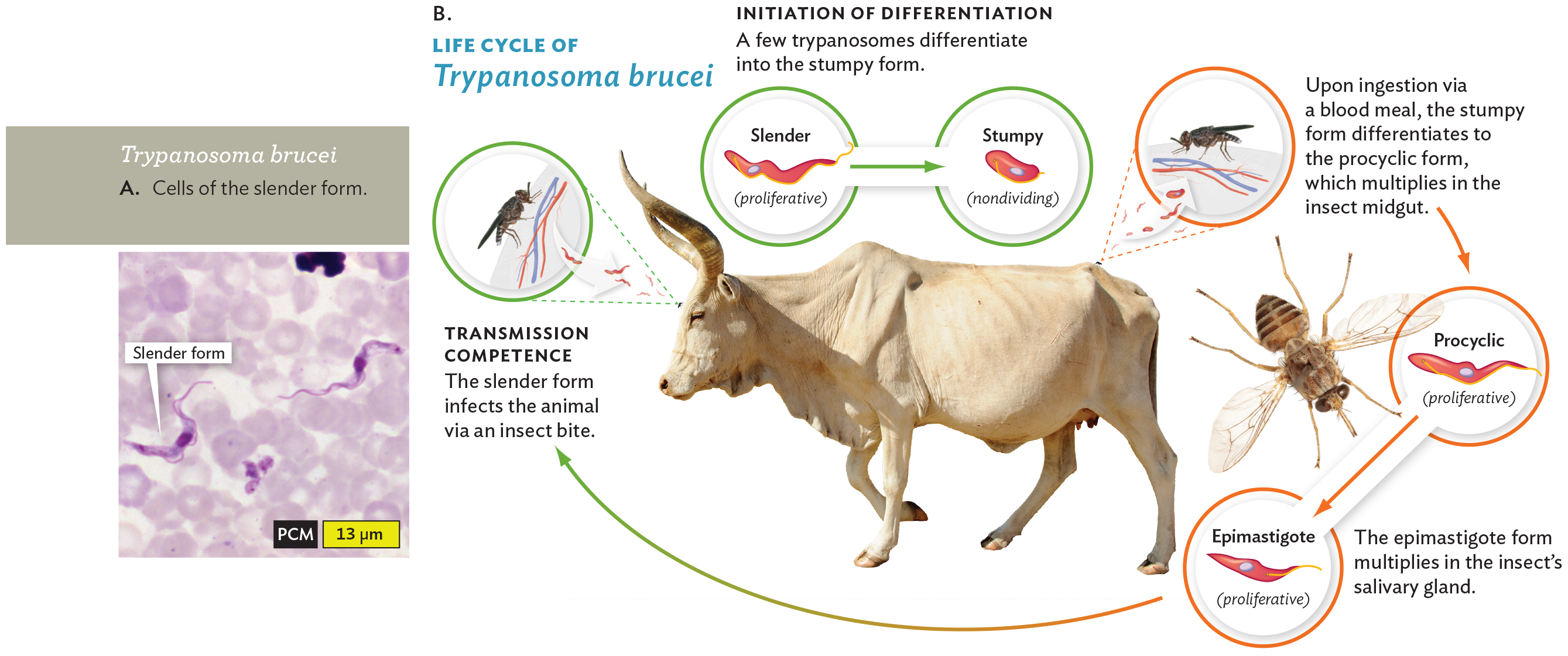
A phase contrast micrograph of Trypanosoma brucei and a diagram of its lifecycle. The first part is the micrograph. It shows a blood smear with the slender form of Trypanosoma brucei. The slender form cells are thin and elongated. They have an appearance similar to snakes. The blood cells are pale staining and disc shaped. The second part is the diagram. There are inset photos of a cow and a tsetse fly at the center of the diagram. First, a tsetse fly releases the slender form of Trypanosoma brucei into the cow when it takes a blood meal. A caption reads, transmission competence. The slender form infects the animal via an insect bite. Within the cow, the slender form is proliferative. It can differentiate into the stumpy form, which is nondividing. A caption reads, initiation of differentiation. A few trypanosomes differentiate into the stumpy form. Another fly takes a blood meal from the cow, this time ingesting a mix of slender and stumpy Trypanosoma brucei cells. A caption reads, upon ingestion via a blood meal, the stumpy form differentiates to the procyclic form, which multiplies in the insect midgut. Within the fly, the procyclic form is proliferative. It multiplies and differentiates into the epimastigote form, which is also proliferative. A caption reads, the epimastigote form multiplies in the insect’s salivary gland. This form is injected into the cow as the slender form when the tsetse fly takes its next blood meal, starting the cycle again.
Trypanosoma brucei, a trypanosome, is carried by the tsetse fly (Figure 11.16B). T. brucei multiplies in the bloodstream of the host animal, causing repeated cycles of proliferation and fever that cause increasing drowsiness, ultimately leading to death, if untreated. This parasite is known for its extraordinary degree of antigenic variation, the ability to generate numerous different versions of its coat protein and thereby evade antibodies of the host immune system. The genome of T. brucei includes 200 different active versions of its variant surface glycoprotein (VSG), the antigen that induces the immune response—as well as 1,600 different “silent” versions that can recombine with “active” VSG to make additional variations. In effect, the parasite overwhelms the host immune system by continually generating new antigenic forms until the host repertoire of antibodies is exhausted.
Diagnosis of trypanosomiasis requires microscopic detection of the parasite in centrifuged blood. The parasite invades many body systems, causing cardiac and kidney dysfunction. In a late stage, the parasites cross the blood-brain barrier, invading the central nervous system. Neurological impairment leads to disruption of the sleep cycle—hence the term “sleeping sickness”—ultimately leading to death.
To infect its human host, T. brucei needs to interconvert among several different forms of its life cycle. Several of these forms are shown in Figure 11.16. For example, the “slender form” (Figure 11.16A) proliferates in the bloodstream of a human or other mammal (definitive host). But the “stumpy form” does not proliferate in the mammal; it needs to be ingested by the tsetse fly (intermediate host), where it lives as the asexual stage. Only the stumpy form can survive the tsetse midgut to develop into the forms that can grow there (procyclic and epimastigote). The salivary gland can transmit the parasite to the next host, where the slender form develops again. Trypanosomiasis is discussed further in Section 21.5.
The molecular basis of conversion among the different developmental forms of a parasite offers targets for new antiparasitic agents. New drugs against T. brucei and malarial parasites are urgently needed to decrease the global impact of these devastating diseases.
Metamonads
Another group of motile parasites is the metamonads. A common example is Giardia intestinalis, the cause of intestinal giardiasis (Figure 11.17). In the United States, Giardia is a frequent nemesis of infant day-care centers, where a significant proportion of caregivers are carriers. Giardia is also endemic in wildlife and contaminates freshwater streams crossed by bears and other animals. The parasite can contaminate community water supplies and is endemic in some Russian cities. Giardia and other metamonads are noted for their anaerobic metabolism and their complete absence of mitochondria, lost through reductive evolution; their metabolism requires the anaerobic intestinal environment of their hosts.
Figure 11.17Giardia intestinalis, a Metamonad Intestinal Parasite
More information
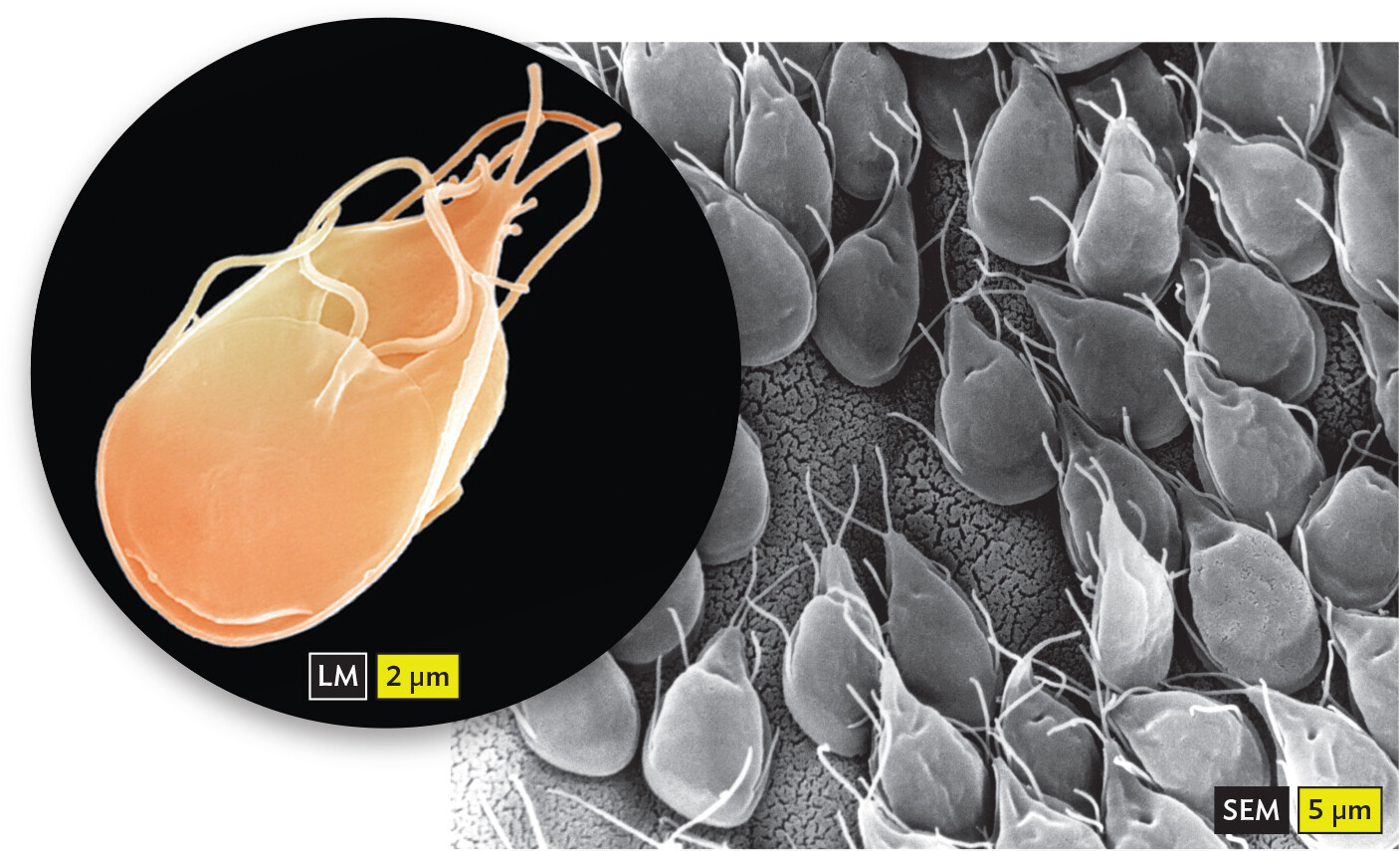
Two micrographs of Giardia intestinalis. The first micrograph shows a single Giardia intestinalis cell. The organism is ovoid shaped with a pointed end. Strands extend from the cell at its pointed end and from a flap at the midpoint of the organism. The second micrograph shows many Giardia intestinalis cells attached to a gerbil intestinal lining. All of the cells attach to the intestine via the flatter, rounded end of the cell body. The pointed ends face the lumen of the intestine.
A.Giardia intestinalis is commonly ingested from mountain streams or transmitted in day-care centers.B. Multiple Giardia cells attached to a gerbil intestinal lining.
Another metamonad is Trichomonas vaginalis, the cause of trichomoniasis, one of the most common sexually transmitted infections (discussed in Chapter 23). In North America, the asymptomatic infection rate among young adults is estimated at 50%. T. vaginalis grows in the urethra and in the vagina. Men rarely report symptoms, but women may experience pain on urination. Untreated infections lead to complications during pregnancy. The infection is treated with the antiparasitic drug metronidazole.
SECTION SUMMARY
Amebas move by using pseudopods. Lobed amebas have large, lobular pseudopods, whereas filamentous amebas have needle-shaped pseudopods.
Free-living amebas are predators. Predatory amebas play an important role in the food web of soil and aquatic communities. They may also harbor pathogenic bacteria.
Entamoeba histolytica causes intestinal amebiasis. The amebas develop into cysts that are expelled in feces and remain dormant for long periods in water.
Naegleria fowleri and Balamuthia mandrillaris cause meningoencephalitis.
Ciliates are predatory protozoa with cilia. Cilia provide motility and generate currents that draw prey into the oral groove.
Apicomplexans are obligate parasites lacking cilia or flagella. Important parasites cause malaria (Plasmodium falciparum) and toxoplasmosis (Toxoplasma gondii). Waterborne opportunistic pathogens include Cryptosporidium parvum and Cyclospora.
Hemoflagellates are obligate parasites that have a single flagellum. Hemoflagellates undergo complex life cycles. They cause sleeping sickness (Trypanosoma brucei), Chagas disease (Trypanosoma cruzi), and leishmaniasis (Leishmania donovani).
Metamonads are parasites with highly degenerate cells. The intestinal parasite Giardia intestinalis causes giardiasis. The urogenital parasite Trichomonas vaginalis causes trichomoniasis.
Thought Question 11.2 How do you think amebas evolved to carry symbiotic bacteria? How do these ameba-carried bacteria endanger humans?
SHOW ANSWERHIDE ANSWER
Amebas engulf bacteria as food. But some bacteria have molecular mechanisms to avoid digestion within the ameba. If the bacteria grow inside the ameba, their respiration may provide nutrients such as vitamins that are useful to the ameba. At the same time, the ameba provides food molecules, and an endosymbiosis may evolve. However, some bacteria, such as Legionella, evolve a partnership with amebas but act as infectious agents within host cells such as macrophages. Inhaled water droplets may contain amebas with intracellular Legionella bacteria, which can cause legionellosis.
Thought Question 11.3 What are the relative advantages and limitations of flagellar motility, ciliate motility, and pseudopodial motility of amebas? Which kind of natural habitat or body environment might favor one kind of motility over another?
SHOW ANSWERHIDE ANSWER
Flagellar motility is the fastest, enabling rapid propulsion of a protist through a dilute watery habitat. However, if the flagellum gets caught or damaged, the cell cannot move. Cilia provide robust movement through a wide range of environments, including places where there are large particles to “dodge.” Cilia also provide a way to generate currents that direct food into the cell’s oral groove. However, cilia may be less effective in a highly viscous medium. Pseudopods enable an ameba to crawl through crevices and highly viscous material. Pseudopodial motility is slow, however, and ineffective in open water.
Thought Question 11.4 Do you think most eukaryotic parasites have genomes that are larger or smaller than those of free-living microbial eukaryotes? Why might parasite genomes be larger—and why might they be smaller?
SHOW ANSWERHIDE ANSWER
Natural selection tends to shrink the genome of a parasite because the smaller the genome, the more progeny can be made from a given host, a limited resource. In addition, the host can provide many complex materials that the parasite need not make, such as amino acids and vitamins. At the same time, however, the parasite needs special mechanisms to attach and manipulate the host and evade the host’s defenses—mechanisms that require extra genes to encode. Consequently, apicomplexans have evolved the apical complex, which penetrates the host cell, and hemoflagellates have evolved a complex series of coat proteins that evade the host’s immune system. Genes encoding these coat proteins expand the size of the parasite’s genome.
1. A tough, protective capsule in tissue that encloses the larva of a parasitic worm within an infected animal. 2. An infectious form of many protozoan parasites, usually passed in feces.
Horizontal gene transmission involving cell-to-cell contact. In bacteria, pili draw together the donor and recipient cell envelopes, and a protein complex transmits DNA across. In ciliated eukaryotes, a conjugation bridge forms between two cells, connecting their cytoplasm and allowing the exchange of micronuclei.
The ability of some protozoan parasites to express multiple variants of their surface protein antigens, thereby overwhelming the host’s ability to produce antibodies against the parasite and enabling it to evade the host immune response.

 Note: Eukaryotic flagella and cilia are very different from bacterial flagella, which are helical protein filaments. Bacterial helical flagella rotate, powered by the proton motive force across the cell membrane.
Note: Eukaryotic flagella and cilia are very different from bacterial flagella, which are helical protein filaments. Bacterial helical flagella rotate, powered by the proton motive force across the cell membrane.