Explain the replication cycle of coronavirus, which has a (+) strand RNA genome.
Describe the structure of the influenza virion.
Explain how the form of the influenza genome enables rapid evolution of novel strains that cause pandemics.
Explain the replication cycle of influenza virus, which has multiple segments of (−) strand RNA.
How does a virus function with a genome of RNA? Many common viruses, such as coronavirus and influenza virus, have genomes of RNA instead of DNA. RNA viruses have especially high mutation rates because their RNA-dependent RNA polymerases lack proofreading.
Coronaviruses such as SARS-CoV-2 have a (+) strand RNA genome that can be translated directly by ribosomes to make proteins (Figure 12.19). On the other hand, influenza viruses have a (−) strand RNA genome that must first be copied to complementary (+) strand RNA to synthesize proteins. Next, we present the replication of coronavirus, followed by influenza virus.
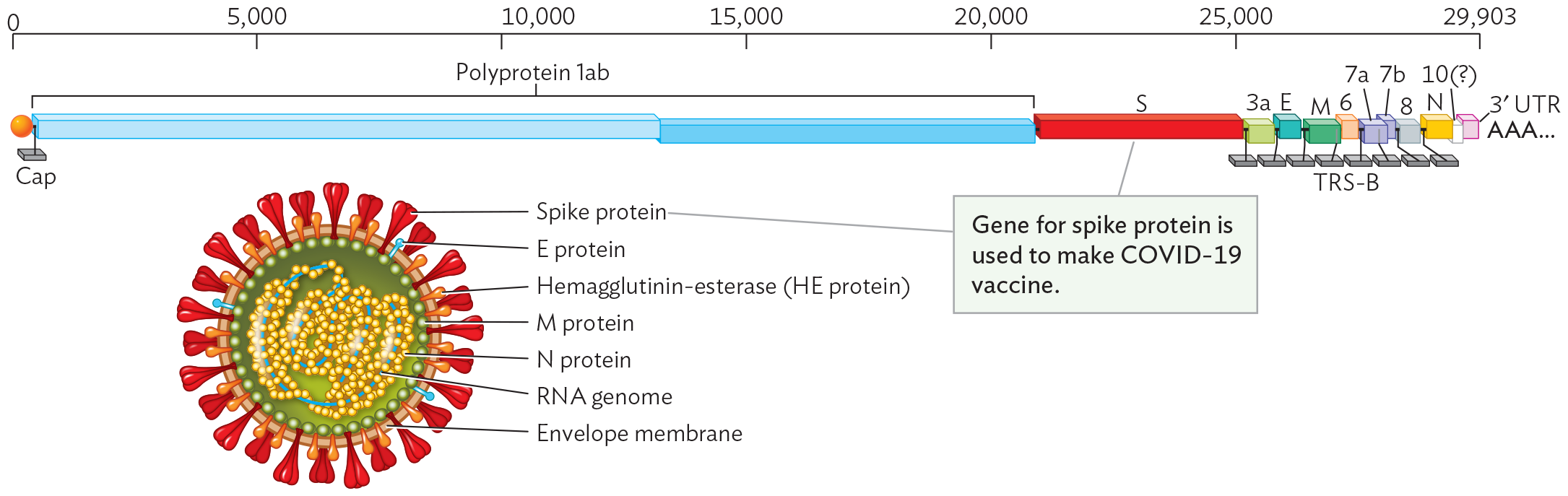
Figure 12.19 SARS-CoV-2 Coronavirus Structure and Genome
More information
A diagram of the S A R S C o V 2 coronavirus structure and genome. The genome is single stranded R N A, as represented by a flat edged block illustration. The genome is 29,903 bases long. The first 20000 bases after the 5 prime cap code for polyprotein 1 a b. The next 5000 bases code for spike proteins. There are a variety of genes between bases 25000 and 29903, ending in the 3 prime A A A tail. An illustration shows the S A R S C o V 2 coronavirus virion. It is roughly spherical. The virion is enclosed within an envelope membrane. The membrane is covered in peg shaped spike proteins, columnar shaped E proteins, and tear drop shaped hemagglutinin esterase or H E protein. The inner surface of the membrane is lined by M proteins. The single stranded R N A genome is packed into the center of the virion. N proteins surround the genome. The caption points to the spike protein in the diagram and reads “Gene for spike protein is used to make COVID - 19 vaccine”. End text.
Genes that encode proteins are shown staggered where translation begins in a different reading frame (triplet start position) from that of the adjacent upstream gene. The (+) strand RNA genome has a eukaryotic cap (7-methylguanosine) attached at the 5′ end (indicated as orange ball). At the 3′ end is a poly-A tail.
SARS-CoV-2 Coronavirus Structure
Many kinds of coronaviruses cause respiratory infections of humans, such as colds. Occasionally, a new strain of coronavirus emerges that causes an epidemic of more severe disease—such as SARS-CoV-2, the cause of the COVID-19 pandemic of 2019–2023. The emerging virus SARS-CoV-2 was notable for its extremely rapid spread, especially within congregate living facilities. “Congregate” describes a situation in which many people live together with a high number of close contacts. To slow the spread of COVID-19, many countries closed schools and in-person businesses. Even so, large numbers of emergency cases swamped hospitals and caused major economic disruption. Although the pandemic alert was ultimately lifted, virus strains continue to circulate, and COVID-19 is now considered an endemic seasonal disease. The pathology of COVID-19 is presented in Chapter 20.
A coronavirus has a core particle of RNA packaged in proteins (nucleoprotein complexes linked by RNA) with a surrounding envelope and spike proteins (Figure 12.19). The appearance of the “crown” of spike proteins in micrographs inspired the name “coronavirus”: corona is Latin for “crown.” The SARS-CoV-2 genome consists of (+) strand RNA (Baltimore Group IV; see Table 12.1). Its 5′ end obtains a cap (7-methylguanosine) from the previous host cell, and the 3′ end receives an added sequence of adenine bases, making it look like normal eukaryotic mRNA. Viral proteins are encoded by various genes. The spike protein (S) is famous as the target of the COVID-19 vaccines and boosters that we now receive to protect ourselves from severe infection. Adaptive immunity and vaccines are presented in Chapter 16.
Other viral proteins include the envelope proteins (E and M) and the nucleocapsid protein (N). Genome replication requires early expression of an RNA-dependent RNA polymerase (replicase), encoded by a portion of the polyprotein 1ab. A polyprotein is a long polypeptide that requires cleavage by a protease enzyme in order to release functional proteins. The viral protease is the target of the inhibitor nirmatrelvir, a component of the antiviral medication Paxlovid, which can decrease severity of COVID-19 illness.
SARS-CoV-2 Coronavirus Replication
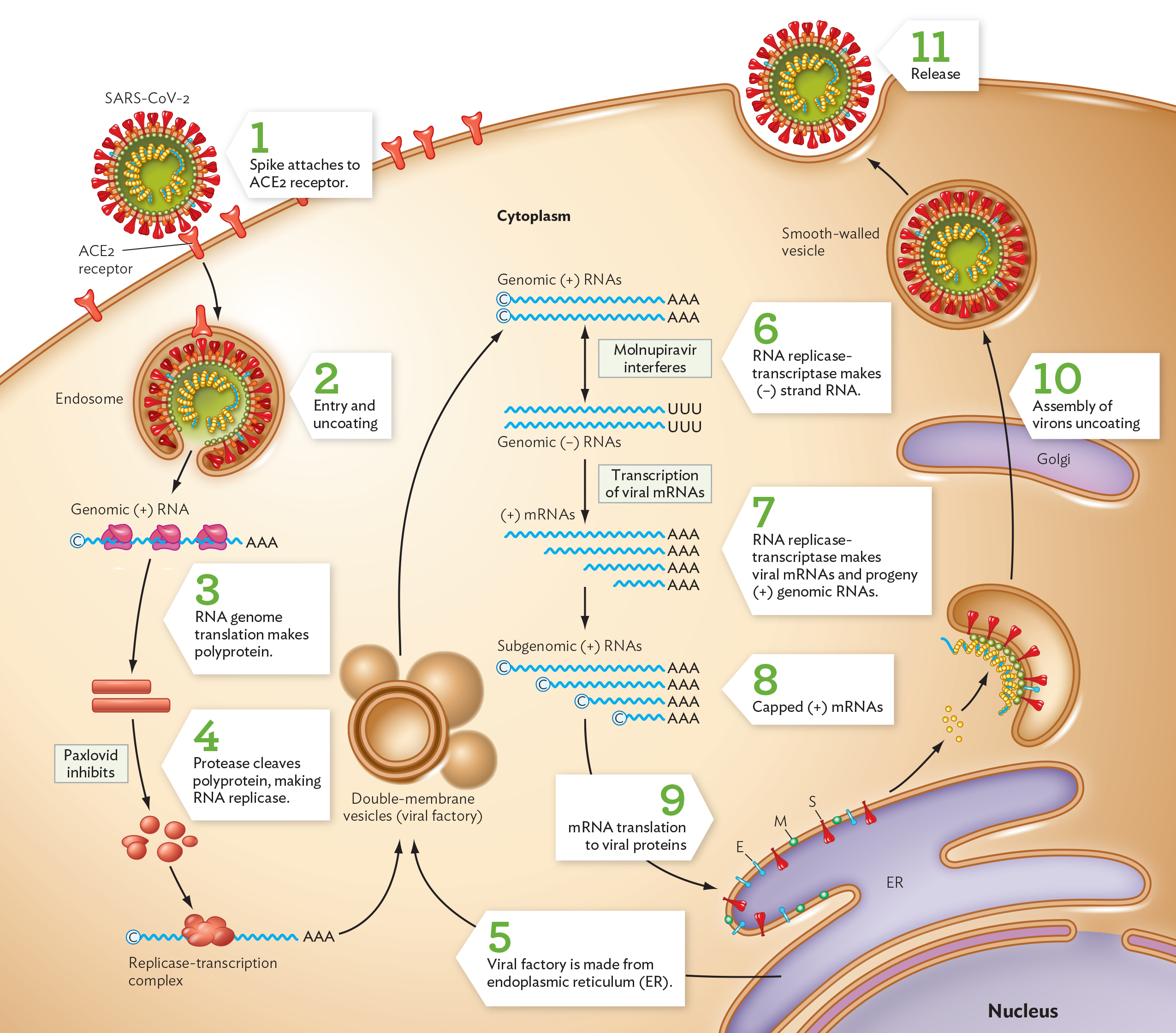
Infection starts when the coronavirus spike protein (S) binds the ACE2 receptor of a host cell (Figure 12.20, step 1). Receptor-spike binding triggers endocytosis of the virion (step 2). Once the infecting virion uncoats in the cytoplasm, its envelope merges with the endosome membrane in a manner that releases the nucleocapsid-coated RNA. The (+) strand viral genome can start translation of its genes as soon as it enters a host cytoplasm (step 3). The gene sequences ORF1a and ORF1b are immediately translated by host ribosomes, forming a polyprotein that will be cleaved by a viral protease (step 4).
A diagram of the S A R S C o V 2 coronavirus replication cycle. There is an illustration of part of a cell. A few key organelles are shown, the nucleus, the endoplasmic reticulum, and the Golgi apparatus. Beginning the cycle, A C E subscript 2 receptor proteins are embedded in the cell membrane. The S A R S C o V 2 virion is spherical. Its outer surface is covered in spike proteins and other membrane proteins. A single stranded R N A genome is packaged within the virion. In step 1, an S A R S C o V 2 spike protein attaches to an A C E 2 receptor in the cell membrane. In step 2, the virion enters the cell via an endosome. Through uncoating, the outer membrane and associated proteins are removed, releasing the S A R S C o V 2 genome into the cell. In step 3, the positive sense R N A genome is translated by host cell ribosomes. Polyprotein is made as a result. In step 4, protease cleaves polyprotein, making R N A replicase. It is noted that Paxlovid inhibits this step. In step 5, the R N A replicase forms a replicase transcription complex that enters into double membrane vesicles, or viral factories. The viral factory is made from host cell endoplasmic reticulum. These vesicles generate genomic positive sense R N A strands. In step 6, R N A replicase transcriptase makes negative sense R N A strands. It is noted that Molnupiravir interferes with this step. In step 7, the genomic negative sense R N A strands are transcribed into viral m R N A. R N A replicase transcriptase makes viral m R N A strands and progeny positive sense genomic R N A strands. In step 8, subgenomic positive sense R N A strands are capped. In step 9, these strands of capped positive sense m R N As are translated into viral proteins. In step 10, these viral proteins are used in the assembly of new virions. This process involves the host E R and Golgi apparatus. In step 11, the newly assembled virions are released into the extracellular space.
Polyprotein cleavage releases subunits of the RNA-dependent RNA polymerase (replicase). All RNA viruses require a special virus-encoded polymerase to make progeny genomes from an RNA template. Most RNA-template polymerases have high rates of mutation compared to DNA polymerases; this is one reason why some kinds of RNA viruses rapidly mutate and generate variant strains so quickly. For example, in 2021 during the COVID-19 pandemic, the Delta variant of SARS-CoV-2 spread rapidly because this mutant virus generated a virus concentration in humans a thousandfold higher than that of the original strain. The subsequent Omicron variant had a large number of mutations and spread even faster.
For further steps of virion construction, a viral factory is formed out of the host endoplasmic reticulum (Figure 12.20, step 5). The SARS-CoV-2 replicase now uses the (+) RNA template to synthesize (−) strand complementary RNA (step 6). The (−) strand RNA then serves as a template to synthesize new (+) strand RNA genomes for progeny virions, as well as mRNA molecules encoding all the viral proteins (step 7). All new RNA molecules are capped at the 5′ end by a host enzyme (step 8). Viral packaging proteins are synthesized (step 9), and all the parts ultimately must be assembled together to make new virions (step 10). The Golgi complex secretes completed virions to the cell surface, and exocytosis then releases the progeny virions outside the cell (step 11).
Culturing Coronavirus
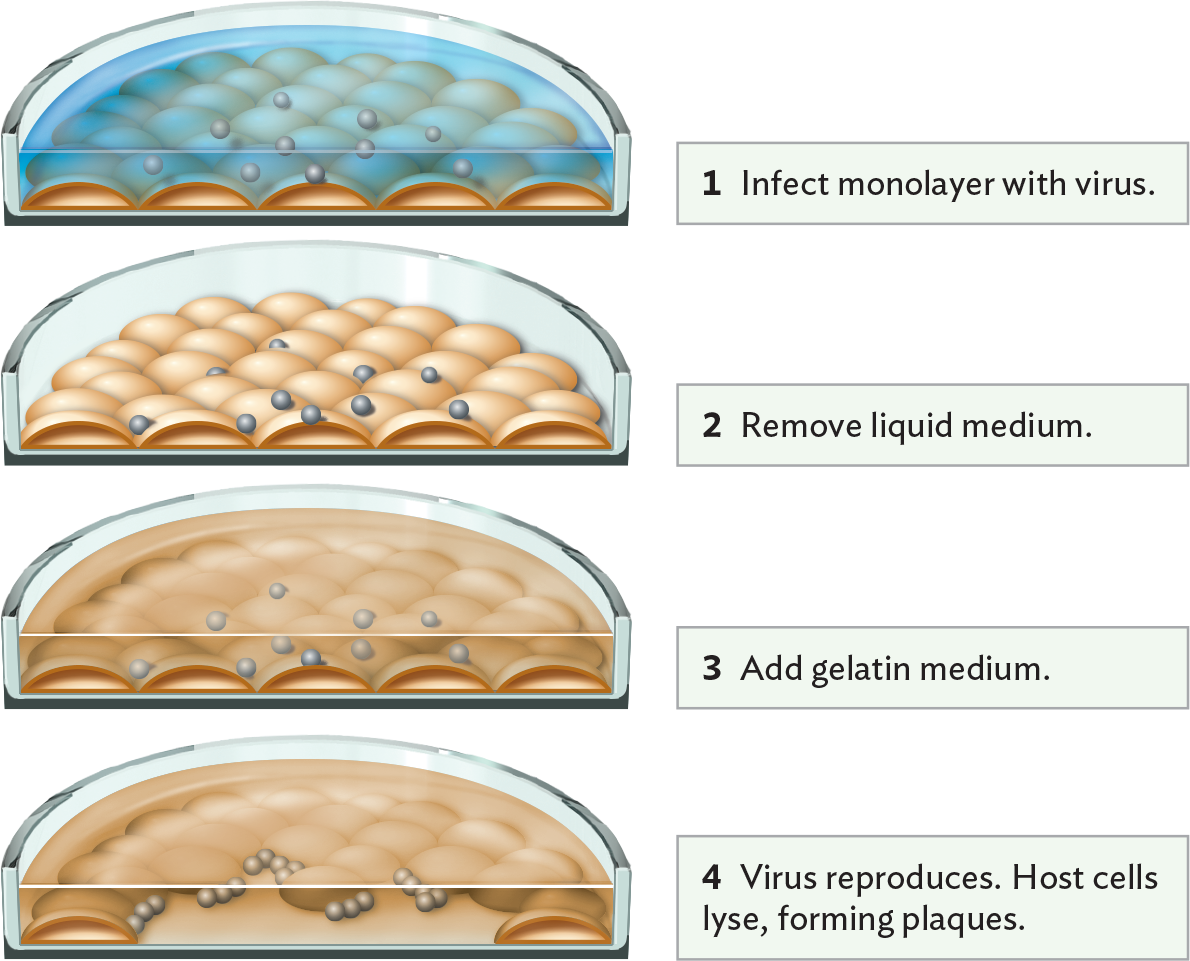
How did researchers learn so much about the coronavirus cycle and devise new antiviral drugs? The virus must be studied in host cell culture. In Section 12.3, we learned how bacteriophages are cultured by plaque assay. For animal viruses, the plaque assay must be modified for host cells in tissue culture (Figure 12.21A). The host cells are usually an immortalized cell line—that is, a line of cells from a cancer in which genetic control of growth is eliminated, so the cells can double in culture indefinitely.
Figure 12.21 Culture of Human Coronavirus
More information
A diagram of a modified plaque assay using cultured host cells. In each step, a cross section view of a petri plate is shown, allowing a view the cells beneath the top layer of the plate. At the bottom of the plate, there is a monolayer of host cells. The cells are dome shaped. In step 1 of the assay, the monolayer is infected with virus by adding a liquid medium containing the virus to the monolayer of host cells. The virus particles are tiny spheres within the liquid medium. In step 2, the liquid medium is removed. The virus particles remain attached to the outer surfaces of cells in the monolayer. In step 3, a gelatin medium is added. This medium covers the monolayer. In step 4, the virus reproduces. Host cells are lysed, which forms plaques. Many of the cells in the monolayer have been destroyed, forming a clear space. Large numbers of virus particles are seen interacting with host cells at the edges of the plaque.
A. Modified plaque assay using cultured host cells. The addition of gelatin medium (step 3) retards the dispersal of progeny virions from infected cells, restricting new infections to neighboring cells. The result is a visible clearing of cells (a plaque) in the monolayer.More information
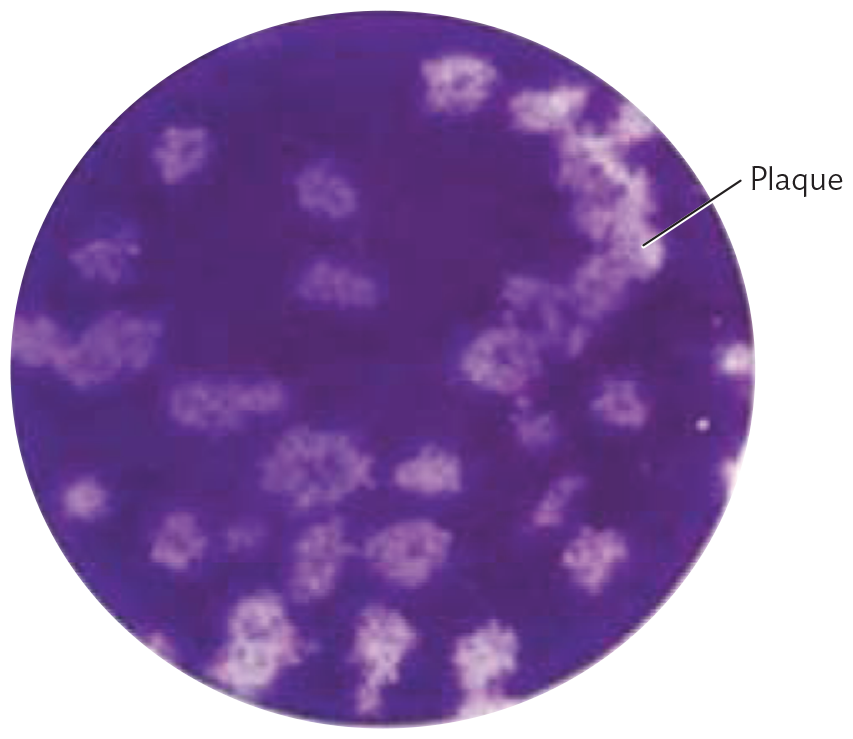
A photo of a plaque assay of human coronavirus H C o V N L 63 plated on colon carcinoma. There is an opaque purple circle with many small irregular clear splotches with fuzzy outlines. The splotches are roughly circular. These are plaques.
B. Plaque assay in which human coronavirus HCoV NL63 suspension was plated on a monolayer of colon carcinoma cells.
Tissue culture involves the growth of human or animal cells in a monolayer on the surface of a dish containing fluid medium. But the fluid medium would quickly disperse any viruses released by lysed cells. To culture viruses, the tissue culture procedure is modified as follows. First, the tissue culture is inoculated with a virus suspension (Figure 12.21A, step 1). Then, after sufficient time to allow for viral attachment to cells, the culture fluid is removed, including any unattached virions (step 2). The fluid is then replaced by a gelatin medium (step 3). The gel retards the dispersal of viruses from infected cells, and as the host cells die, plaques are observed (step 4). Figure 12.21B shows the appearance of plaques of human coronavirus (HCoV NL63) cultured on colon carcinoma cells. This coronavirus causes mild respiratory symptoms; it is related to SARS-CoV-2.
CASE HISTORY 12.2
Seasonal Surprise: Influenza at College
More information
A photo of a woman wearing an N 95 face mask. The face mask is securely fitted to her face, leaving no gaps between the mask and her skin. The mask fits from the bridge of her nose to down below her chin. Straps around the back of her head and neck keep the mask in place.
Eighteen-year-old Sofia was a first-year student at a small private college in Ohio. In the fall of 2022, she was excited to start college at a time when the COVID-19 pandemic restrictions had largely been lifted and most students no longer wore masks. She lived in a residence hall with 150 other students, which is considered a congregate living facility. By November, she was staying up late many nights to study for exams and complete papers. Her stress levels and sleep deprivation decreased her body’s innate immunity, the ability to fight off illness (see Chapter 15).
One night, Sofia felt hot, flushed, and unsteady on her feet. Her thermometer showed a temperature of 39°C (102°F). She started coughing hard, and she felt extremely tired. She took a home test for COVID-19, but the result was negative. A health worker wearing a face mask came to Sofia’s room to administer a test for influenza A, which eventually showed positive. Many other students were experiencing influenza at the same time.
Sofia was confined to her room; her roommate decided to stay with a friend. Sofia lay in a bed and tried to sleep. In the morning, a nurse’s aide left a tray of food and medication, including oral oseltamivir (Tamiflu) and inhaled zanamivir (Relenza), outside Sofia’s room. Sofia had to drag herself out of bed to open the door and get the tray; she tried to drink as much water and soup as possible. She was required to stay in her room until 2 days after her temperature returned to normal. Sofia had a lot of work to make up once the illness had passed, and she required an incomplete grade for some courses so she could finish her assignments during winter break. She and most other infected students made a full recovery, although one student was hospitalized with life-threatening complications.
Influenza A virus is an orthomyxovirus (Baltimore Group V; see Table 12.1). Influenza is one of the most common life-threatening viruses in the United States. The virus infects cells of the upper respiratory mucosa, causing fever, sore throat, headache, cough, and shortness of breath. Each year, influenza A infects approximately 10% of the US population, causing about 36,000 deaths annually. The elderly are most susceptible, but in some years a new strain increases mortality among young people.
Influenza strains show antigenic drift (see Section 12.2) in that their envelope proteins continually mutate, evading the host immune system; as a result, new strains emerge annually—the “seasonal flu.” To avoid seasonal influenza, an annual vaccination is recommended. Beyond the annual variant strains, at wider intervals extremely virulent strains of influenza emerge that cause pandemic mortality. For example, the famous influenza pandemic of 1918 infected 20% of the world’s population and killed more people than World War I. The 1918 strain arose as a mutant form of an influenza strain infecting birds. Such extremely virulent strains usually arise through antigenic shift, in which genes reassort from two or more different influenza viruses. Antigenic shift represents a larger change than antigenic drift, which produces the smaller mutations observed in seasonal influenza strains. Today, other “avian influenza” strains continue to emerge by viral reassortment in animal hosts. As of this writing, however, the known avian strains have limited tropism for the human respiratory tract, and none have mutated to a form readily transmitted between humans (discussed further in Chapter 20).
Antigenic shifts and drifts enable viruses to evade the host immune system. Influenza type A is the most capable of undergoing genetic change, so it more commonly leads to seasonal disease. Type A influenza is classified into subtypes based on the kind of hemagglutinin (H) and neuraminidase (N) expressed on the viral surface, giving rise to some very well-known strains, such as H1N1 (swine flu) and H5N1 (avian flu).
Influenza Virus Structure
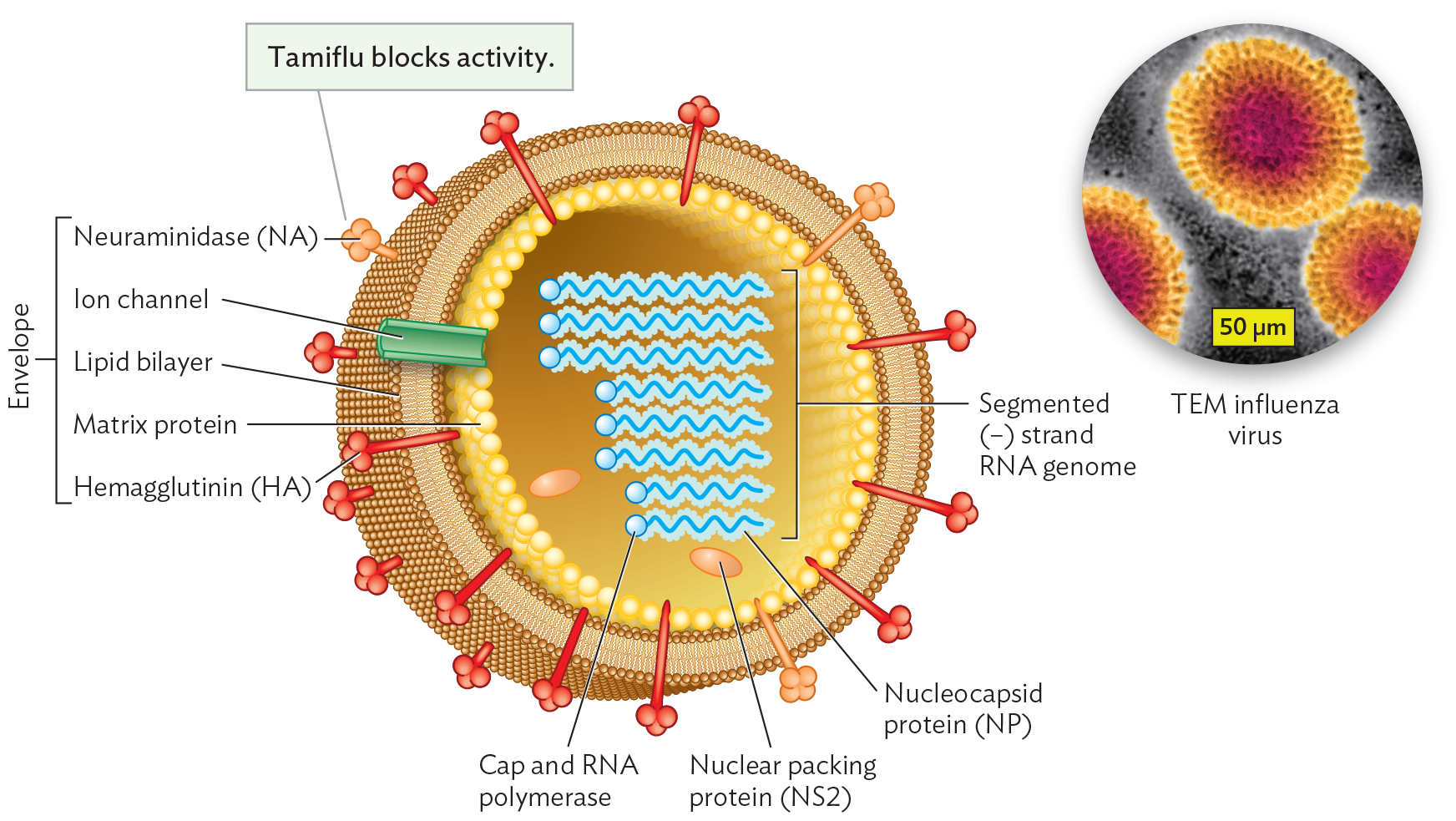
How do major pandemic strains of influenza arise? The cause can be understood from the structure of the influenza virion and its mode of replication. The influenza virion is asymmetrical; it has no fixed capsid (Figure 12.22). Unlike the coronavirus genome, the influenza genome consists of (−) strand RNA.
Figure 12.22 Structure of Influenza Virus
More information
An illustration of the structure of Influenza virus with an inset micrograph of influenza virus particles. The illustration shows a spherical virion. The envelope enclosing the virion consists of a lipid bilayer. Hemagglutinin or H A and neuraminidase or N A proteins are embedded in the lipid bilayer. There are also a few ion channels in the envelope. Matrix proteins form the inner lining of the envelope. At the center of the virion is the segmented negative sense strand R N A genome. Each strand includes a cap and an R N A polymerase as well as a nucleocapsid protein. There are also a few nuclear packing proteins within the center of the virion. The inset transmission electron micrograph is centered on an influenza virus particle. The virion is roughly spherical. It has a distinct envelope with a high volume of embedded proteins. The interior is darker and grainy where the genome is packed.
Diagram of influenza virion structure, showing the envelope (brown), envelope proteins (HA and NA), matrix protein (yellow), RNA segments with attached polymerase, and nuclear packing protein NS2. Inset: An influenza A virion. The brush-like border coating the envelope consists of glycoproteins, hemagglutinin (HA), and neuraminidase (NA). Envelope proteins involved in receptor binding (HA and NA) are also called spike proteins.
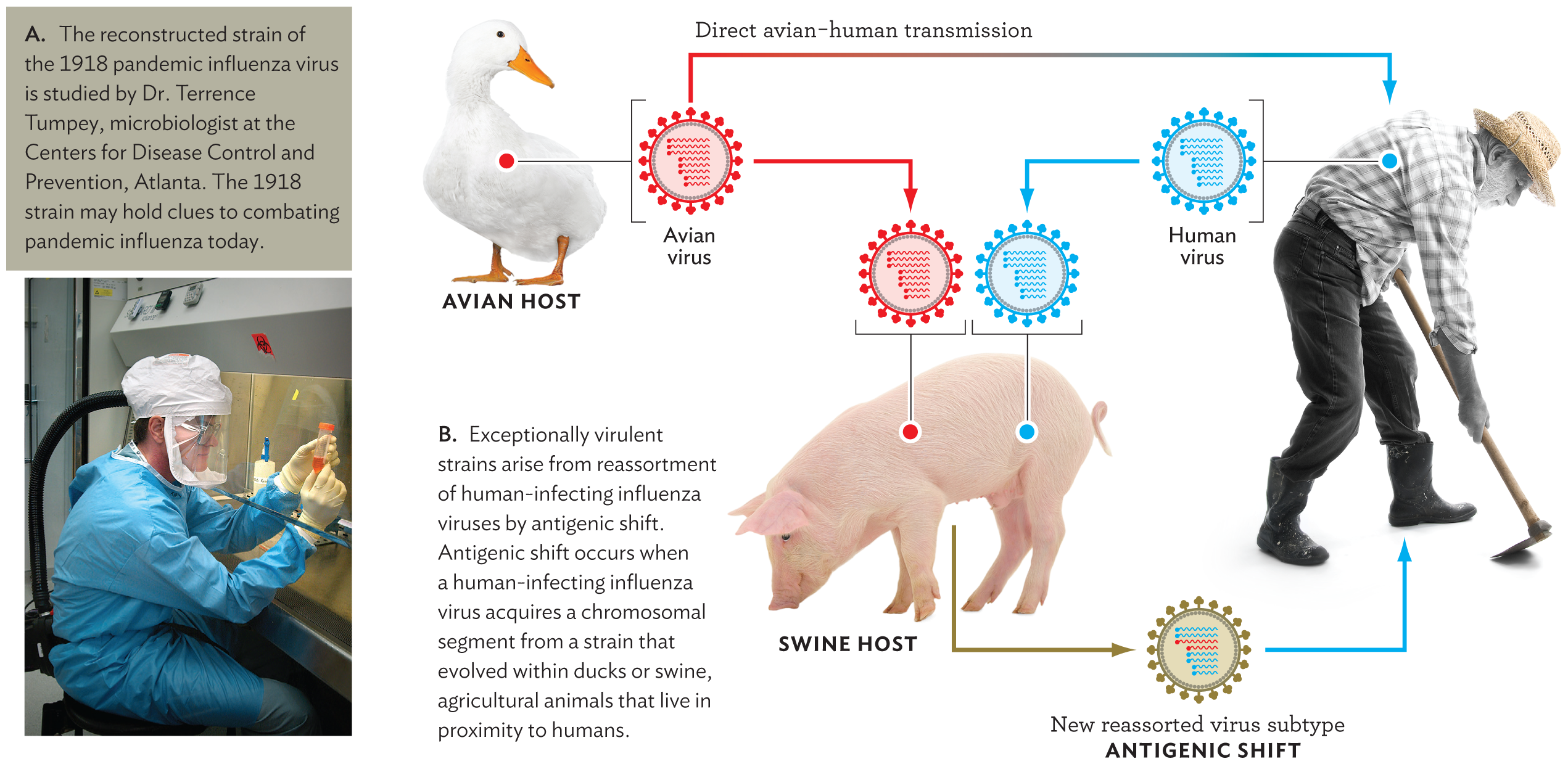
An unusual feature of the influenza genome is that its genomic RNA is divided among eight chromosome segments. Each segment contains a different essential gene. When influenza virus infects a cell, the eight RNA segments can reassort as virions combining segments from a coinfecting virus of a different strain. Human influenza RNA segments can reassort even with segments from a strain that normally infects a different host species and cannot complete its replication in the human cell (Figure 12.23). Such reassortment is particularly likely with influenza strains from birds and pigs, which are often raised in close proximity to humans. The reassortment may generate a new strain able to infect humans but possessing novel gene sequences unfamiliar to the host immune system. This interspecies reassortment is what leads to the most virulent pandemic strains of influenza.
Figure 12.23 Reassortment between Human and Avian Strains Generates Exceptionally Virulent Influenza Strains
More information
A photo of Dr. Terrence Tumpey and a schematic explaining the reassortment of human and avian influenza virus strains. The first part is the photo. Dr. Tumpey is working in a biological safety cabinet. He is wearing full protective gear and a respirator. The second part is the schematic. There is an avian host, a swine host, and a human host. An avian virus from the avian host and a human virus from the human host are both able to replicate within a swine host. This leads to a new reassorted virus subtype that can infect the human host. This is called antigenic shift.
Within the influenza virion, the RNA segments are coated by nucleocapsid protein (NP). Each NP-coated RNA segment also possesses a bound RNA-dependent RNA polymerase complex, poised for synthesis. The prepackaged polymerase is necessary because, during infection, each (−) strand RNA segment must be transcribed to a (+) strand mRNA for translation—but the host has no RNA-dependent RNA polymerase. So the virus needs to bring its own, a copy of which is all set to go on each segment.
The virion’s RNA segments are loosely contained by a shell of matrix proteins. The envelope originates from the phospholipid membrane of the host cell. The membrane incorporates viral proteins, such as hemagglutinin (HA) and neuraminidase (NA), that peg the membrane to the matrix, maintaining an intact but flexible structure. The HA and NA proteins differ slightly in sequence among different strains. The sequence differences of the HA and NA proteins give rise to the H and N numbers, such as in strain H1N1 (the so-called swine flu) and H5N1 (an “avian flu” strain). Neuraminidase also acts as an enzyme whose activity can be blocked by the antiviral agent oseltamivir (Tamiflu), one of the main drugs available to treat influenza.
Given that influenza infection requires all eight RNA segments, the viral assembly process offers opportunities for “mistakes” that lead to new infectious strains. Suppose a host cell gets infected by two different influenza strains, such as an avian strain and a human strain. By “mistake,” an avian RNA segment could combine with human segments, forming a reassorted virus. The reassorted virus could generate avian-like proteins that would be invisible to the human immune system, causing antigenic shift and potentially resulting in an exceptionally virulent strain.
Influenza Replication
What determines how influenza spreads—and how can we stop it? As we will discuss in Chapter 20, surprisingly small details of the host receptor structure may determine whether a strain of influenza virus will spread directly between humans. Accordingly, avian influenza strain H5N1 is rarely transmitted between humans. However, a mutation in the HA gene could allow the envelope protein to bind human cells in the upper respiratory tract, which would increase transmission and possibly lead to an influenza pandemic. Much research addresses hemagglutinin (HA) and other viral proteins as targets of possible drugs.
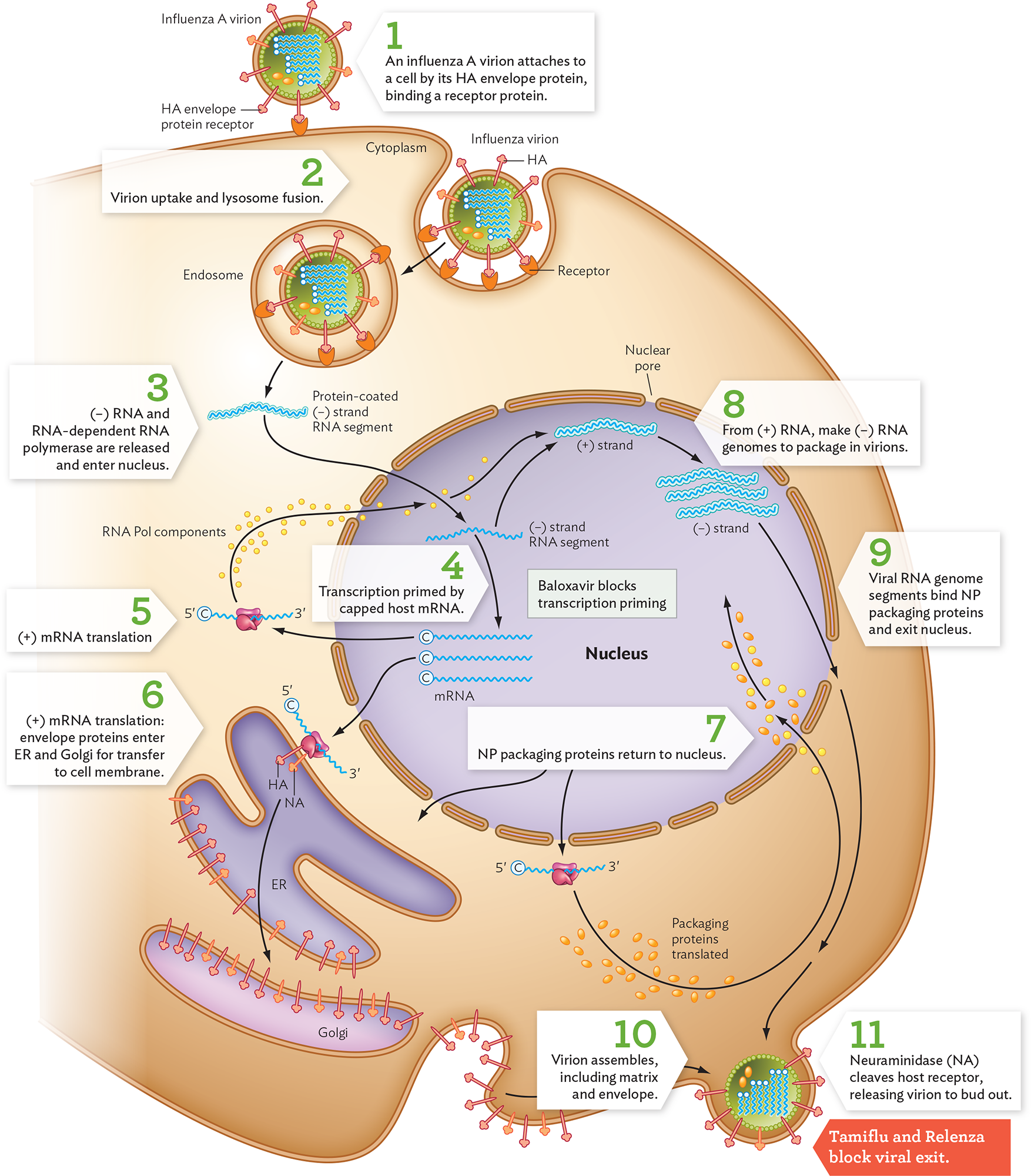
An influenza virion attaches to a cell when its HA envelope protein binds to a host cell receptor, a glycoprotein (Figure 12.24, step 1). After the influenza virion undergoes endocytosis, the endocytic vesicle fuses with a lysosome (step 2). The low pH of the lysosome contents causes hydrogen ions to leak through the virion’s ion channel, an important drug target. The hydrogen ions (low pH) cause the virion to disassemble and fuse with the endocytic membrane. As the membranes fuse, the virion contents are released into the cytoplasm, including all the viral (−) RNA segments with their prepackaged polymerases (step 3). The RNA segments pass through nuclear pore complexes into the nucleus. Within the nucleus, the viral RNA-dependent RNA polymerases that are attached to each (−) RNA synthesize (+) RNA for mRNA (step 4). Viral transcription is primed by pieces of capped host mRNA that are cleaved by a viral enzyme and attached to the 5' end of viral mRNA. This process is the target of the antiviral drug baloxavir marboxil. The capped viral mRNA then returns to the cytoplasm for translation by host cell ribosomes (step 5).
Figure 12.24 Influenza Virus Replication Cycle
More information
A diagram of the influenza virus replication cycle. There is an illustration of the host cell, with a few key organelles inside: the nucleus, the endoplasmic reticulum, and the Golgi apparatus. Receptor proteins are embedded in the cell membrane. In step 1, an influenza A virion attaches to the cell by its H A envelope protein by binding to a host cell receptor protein. The virion is spherical. N A and H A proteins are embedded in its envelope. The single stranded, negative sense R N A genome is located in the center of the virion. In step 2, the virion is taken into the cell and lysosome fusion occurs. The virion is taken deeper into the cell within an endosome. In step 3, negative sense R N A and R N A dependent R N A polymerase are released into the cytoplasm. These enter the cell nucleus. The strand is protein coated before it enters the nucleus. In step 4, transcription is primed by capped host m R N A. It is noted that Baloxavir blocks transcription priming. In step 5, the positive sense capped m R N A strands leave the nucleus and are translated by host ribosomes. R N A polymerase components are produced, which travel back into the nucleus. In step 6, further positive m R N A strand translation takes place. Envelope proteins enter the endoplasmic reticulum and Golgi apparatus for transfer to the cell membrane. In step 7, N P packaging proteins are translated and return to the nucleus. In step 8, the negative sense R N A genome is made from the positive sense R N A strands in the nucleus. In step 9, the negative sense viral R N A genome segments bind N P packaging proteins and exit the nucleus. In step 10, the virions are assembled at the host cell membrane. Assembly includes the matrix and envelope. In step 11, neuraminidase or N A, cleaves the host receptor, releasing virions to bud out. It is noted that Tamiflu and Relenza block viral exit.
HA, hemagglutinin; NP, nucleocapsid proteins; ER, endoplasmic reticulum.
Animation: Influenza Virus Replication
Some of the mRNA molecules encode envelope proteins, which are made by ribosomes attached to the endoplasmic reticulum (ER; Figure 12.24, step 6). The ER transports the envelope proteins to the Golgi and ultimately to the cell membrane, where they will coat progeny virions. The NP proteins needed to package the viral RNA (packaging proteins) must return to the nucleus (step 7). In the nucleus, the (+) RNA strands now serve as templates for RNA-dependent RNA polymerase to make complementary (−) RNA segments for the progeny viral genomes (step 8). The (−) RNA viral genome segments become coated with proteins (step 9). At last, the protein-coated (−) RNA segments exit the nucleus and are transported to the cell membrane for packaging into a loose capsid of M1 matrix proteins (step 10). As they assemble, the capsids acquire envelope membrane from the host, incorporating viral envelope proteins. The mature virions now bud out (step 11) in a massive release of virions that destroys the host cell.
The final budding step requires the action of neuraminidase (NA), an enzyme embedded in the envelope of the progeny virions; the enzyme cleaves host surface sugar molecules that bind exiting virions to the cell surface. NA is the target of oseltamivir (Tamiflu) and zanamivir (Relenza)—at present the most effective drugs for decreasing the symptoms of influenza.
Compared with the papillomavirus cycle, the replication cycle of influenza virus is very rapid, causing swift destruction of mucosal cells. But the host immune response is also swift. If successful, the host immune response completely eliminates the flu virus within 1 or 2 weeks. Unlike for HPV, there is no long-term persistence in the host cells. So influenza virus is committed to rapid infection and highly efficient host-to-host transmission—as demonstrated during the 2009 H1N1 outbreak.
In the next section we examine one more virus, with another very different infection strategy: the retrovirus HIV, the cause of AIDS. Like influenza virus, HIV has an RNA genome with a high mutation rate. But the HIV virion generates a DNA copy of its genome that, like the human papillomavirus genome, can integrate into the host genome.
SECTION SUMMARY
Coronaviruses are (+) strand RNA viruses that cause respiratory disease. The SARS-CoV-2 coronavirus is the cause of COVID-19 disease, which led to the pandemic of 2019–2023 and is now an endemic seasonal virus.
Coronavirus replication involves RNA-dependent RNA polymerase and formation of a polyprotein.Enzymes such as RNA-dependent RNA polymerase and polyprotein protease are targets of antiviral medications.
Influenza virus causes periodic pandemics of respiratory disease. New virulent strains arise through reassortment of viral genome segments from human, avian, and swine strains.
Antigenic shift is the reassortment of genome segments from a viral strain that infects one species with segments from a strain that infects a different species. Antigenic shift can lead to highly virulent strains of a virus that may cause widespread outbreaks of infection.
Influenza virus consists of segmented (−) strand RNA. Each segment is packaged with nucleocapsid proteins. Segments from different strains reassort by coinfection of a host cell.
Nucleocapsid and matrix proteins enclose the RNA segments of influenza virus. The matrix is enclosed by an envelope containing spike envelope proteins HA and NA.
The influenza virion is internalized by endocytosis, followed by lysosome fusion. Low pH triggers viral envelope fusion with the endosome membrane, releasing the viral contents into the cytoplasm.
Viral (−) RNA segments attached to RNA-dependent RNA polymerase enter the nucleus. The RNA-dependent RNA polymerase makes new (+) sense and (−) sense strands. The (+) sense viral mRNAs return to the cytoplasm for translation, while (−) sense progeny genome segments are packaged into virions.
Influenza virions are assembled at the cell membrane. The capsid, matrix, and (−) strand RNA components are packaged and enclosed by host cell membrane that contains viral envelope proteins. Completed virions bud out and destroy the host cell.
Thought Question 12.6 Researchers in Hong Kong are conducting a regular survey of the genomes of influenza strains emerging in a particular swine-processing facility, where swine are shipped from many different regions. What might the researchers learn that would be important for human public health?
SHOW ANSWERHIDE ANSWER
Swine are known to be an important source of reassorted strains of influenza virus that can cause human pandemics. For example, an avian strain and a human strain can coinfect swine and generate a new strain that infects humans. The infectivity of influenza strains depends on the sequence and binding properties of their envelope proteins. Researchers can study the envelope proteins of influenza strains appearing in swine and predict whether these strains may “escape” into human populations. If a dangerous strain is found, it might be possible to devise a vaccine ahead of an outbreak.
A major change in a viral antigen that occurs when two (or more) strains of a virus with a segmented genome infect the same host cell and reassort their genome segments. The resulting chimeric virus can express a new combination of surface antigens (such as influenza hemagglutinin and neuraminidase) that can alter virulence or enable the infection of new hosts.