Describe differences between “normal” microbiota and pathogens.
Discuss the relationship between infection and disease and between virulence and pathogenicity.
Differentiate between infectious dose and lethal dose.
Discuss the fundamental attributes of a successful pathogen.
Are we alone? You may not want to think about it, but we harbor at least as many microbial hitchhikers as we have human cells in our body. The collection of bacteria, archaea, and eukaryotic microbes that call us home is known as our normal microbiota or microbiome. (We will discuss this more deeply in Chapter 14.) Although microbiota are commonly called “commensal organisms” (organisms that take sustenance from their host but do not give it), most members of our microbiome derive benefit from and give benefit to their hosts; this give-and-take defines mutualism, or a mutualistic relationship.
To stay with their host, normal microbiota have proteins called adhesins on their surface that enable them to attach to and colonize epithelial cells lining mucous membranes (intestine, urinary tract, mouth, nose). Colonization is the ability of a microbe to stay attached to a body surface and replicate (Figure 2.1). Think about your own intestine. Without an ability to attach, some very useful bacteria would too easily be swept out of your body along with feces or urine. Nevertheless, the composition of an individual’s microbiome can fluctuate, as different bacteria and strains of microbial species enter and leave our bodies depending on what we eat, how we exercise, and whom we meet. A changing microbiome can alter our susceptibility to pathogens, strengthen or weaken our immune system, and—believe it or not—influence our weight and mental health.
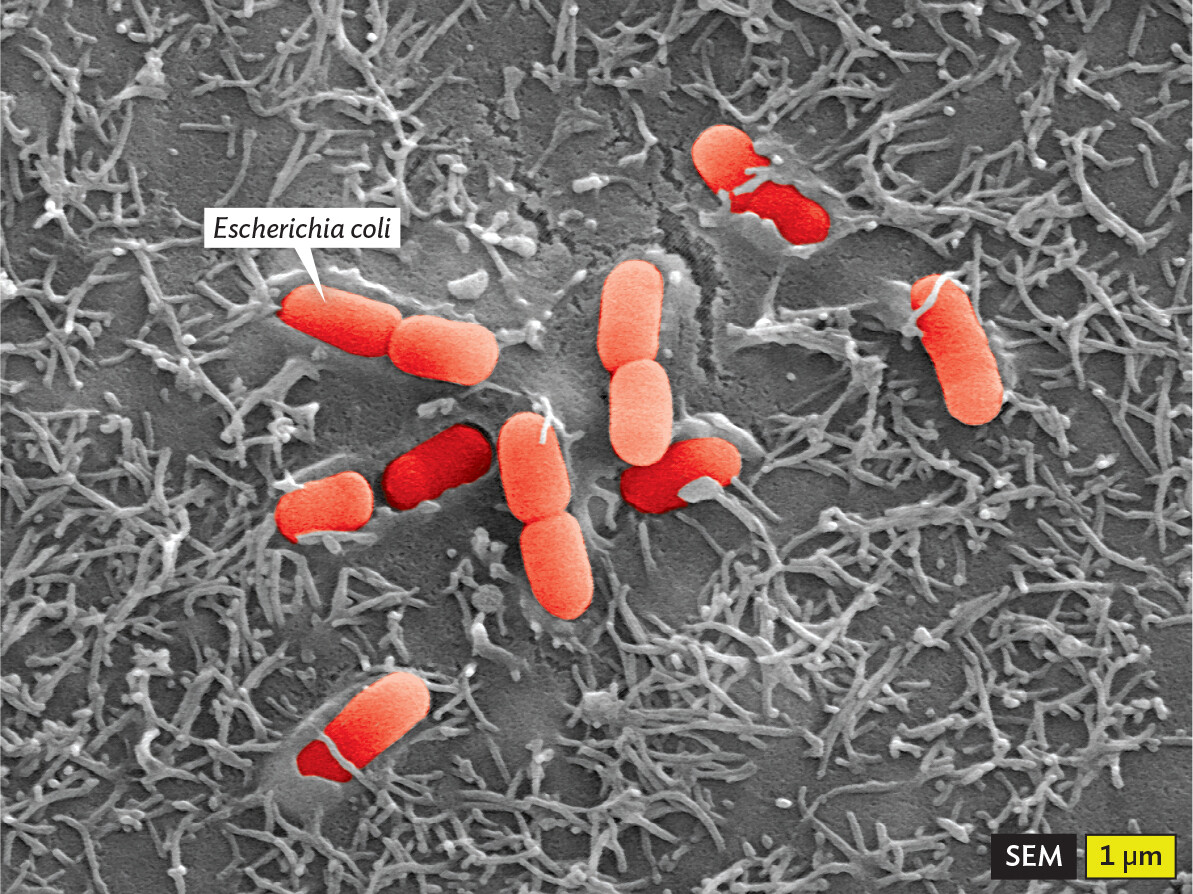
Figure 2.1 Bacterial Attachment to Human Cells
More information
A scanning electron micrograph of Escherichia coli cells adhering to human cells. Each bacterium is about 1.5 micrometers in length and 0.5 micrometer in width. The bacteria adhere to the human cells by sticking to hair like appendages on the outer surfaces of the cells. The E coli cells have been colorized red. The human cells are in greyscale.
3D micrograph showing a small cluster of bacteria (Escherichia coli) adhering tightly to human cells. Note that these bacteria are rod-shaped. Some are shown dividing. The image is artificially colorized.
Defining which species are members of our normal microbiome can be tricky. Take, for example, the bacterium Escherichia coli. E. coli is a part of our normal intestinal microbiome. We all harbor this organism. But news outlets carry alarming stories about E. coli causing death and disease. So does E. coli cause disease or doesn’t it? The answer is yes—to both questions. Every species of bacteria, including E. coli, is composed of different strains that display unique characteristics. Some strains of E. coli contribute to our health as residents of the colon and are a natural part of our microbiome. Other strains, however, can cause diarrhea, septicemia (blood infection), meningitis, and urinary tract infections.
Those harmful strains ofE. coli are, by definition, pathogens. A pathogen is any bacterium, virus, fungus, protozoan (single-celled eukaryotic organism), or worm (helminth) that causes disease. However, defining just what makes a pathogen a pathogen is not so easy. The pathogenicity of an organism—meaning its ability to cause disease—depends on a combination of the organism’s genetic makeup, its location in or on the host’s body, and the effectiveness of the host’s immune response. For example, an E.coli strain that is innocuous in you could possibly kill your immunocompromised neighbor.
The term “pathogen” is typically used by health professionals to refer only to bacterial, viral, and fungal agents of disease, so what do they call other eukaryotic infectious agents such as protozoa and worms? In medical usage, the word “parasite” is reserved for these types of disease-causing agents. However, in broad biological usage, parasite is defined to include any organism that colonizes and harms its host.
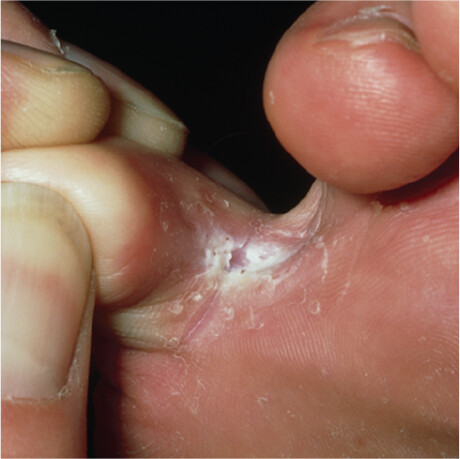
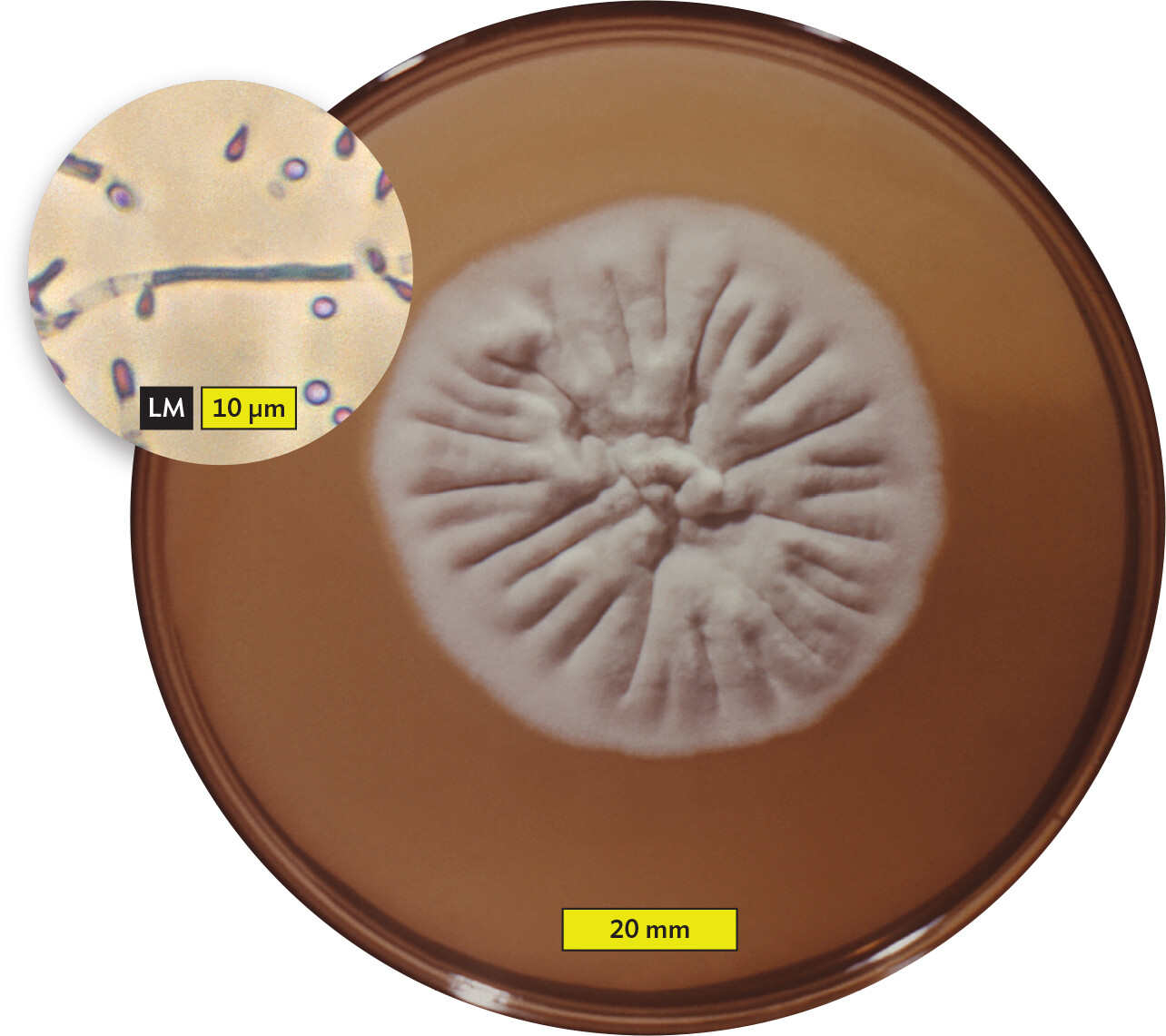
Pathogens and parasites infect their animal and plant hosts in different ways and enter into different host-pathogen relationships, depending on the site of colonization and the capabilities of the organism. For instance, some organisms can live only on the surface of a host; an example is the parasitic fungus Trichophyton rubrum, one cause of athlete’s foot, also called tinea pedis (Figure 2.2A). T. rubrum, which grows on solid media as filaments called hyphae (Figure 2.2B; see Section 11.2), does not have the molecular tools needed to grow inside a host. Insects such as fleas or lice that live on the body surface are also called parasites—ectoparasites. In contrast, Wuchereria bancrofti, the parasitic worm that causes elephantiasis, is an endoparasite: it causes disease by living inside the body (Figure 2.3). Although W. bancrofti is not a problem in the United States, it is found throughout central Africa, Asia, and New Zealand.
Figure 2.2A Skin Fungus
More information
A photo of an athlete’s foot infection in the crease between two toes. There is a white and flaky to powdery growth on the skin in this region. The surrounding skin is red and irritated.
A. Athlete’s foot is caused by the fungus Trichophyton rubrum.More information
A photo and light micrograph of Trichophyton rubrum. A photo of a Trichophyton rubrum colony growing on an agar plate. The colony has a round cottony surface and is white to cream colored. The outer edge has a fuzzy appearance. The central region of the colony is wrinkled, with lines extending from the circumference to the center. The colony is about 60 millimeters in diameter. A light micrograph of Trichophyton rubrum hyphae and spores. The hyphae are thin filamentous strands with multiple segments. The spores are spherical and float in the area around a visible hypha. The hypha is about 20 micrometers in length and 1 micrometer in width. The spores are each about 2 micrometers in diameter.
B.T. rubrum colony shape. Inset is a magnification of T. rubrum showing the branching filaments (called hyphae) with small spores (called microconidia) that form the colony.
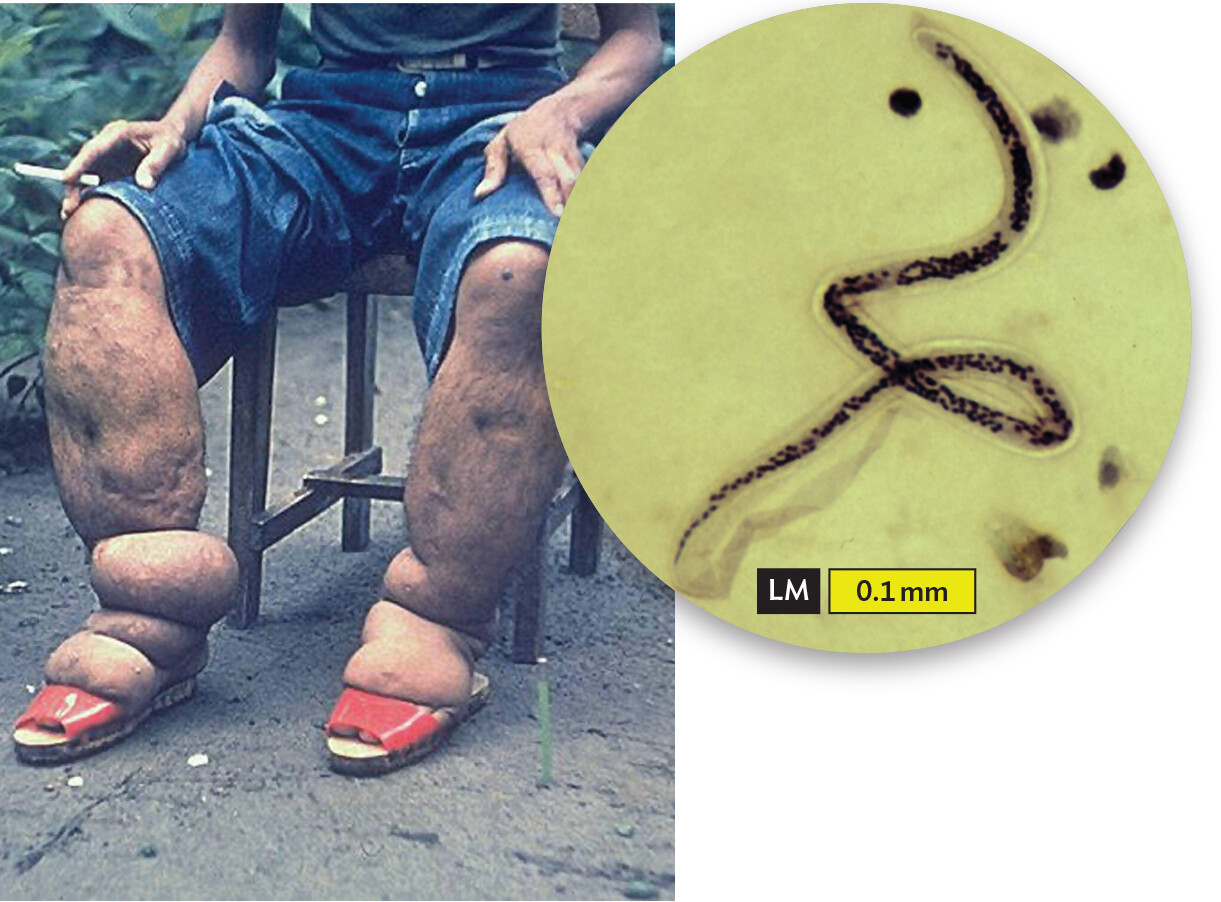
Figure 2.3 An Endoparasite
More information
A photo of a patient afflicted with the parasite Wuchereria bancrofti with an inset light micrograph of the parasite. The photo shows the legs of a man with elephantiasis. His lower legs are severely swollen and pitted. The inset micrograph shows a young worm. It is long and thin with tapered ends.
The disease filariasis, commonly known as “elephantiasis” because of the appearance of the affected limbs, is caused by the worm Wuchereria bancrofti (inset), which enters the lymphatics (vessels connecting lymph nodes) and blocks lymphatic circulation. Adult worms are threadlike and measure 4–10 cm (1.5–4 inches) in length. The young microfilaria shown are approximately 0.5 mm in length.
Note: The various types of microscopy used to see microbes will be described in Chapter 3. For now, let’s focus on appreciating the appearance and size of these organisms. For example, in the inset photo in Figure 2.3, visually compare the magnified W. bancrofti with the length of the scale bar.
The Lingo of Pathogenesis
Before we talk about how microbes cause disease, it is helpful to know the terminology of pathogenesis. An infection occurs when a pathogen or parasite enters or begins to grow on a host. The term “infection,” however, does not necessarily imply overt disease, in which a patient develops symptoms. Any potential pathogen growing in or on a host is said to cause an infection, but that infection may be temporary and undetected if immune defenses kill the pathogen before symptoms develop. Indeed, most infections go unnoticed. For example, every time you have your teeth cleaned by a dentist, your gums bleed and your oral microbiota transiently enter the bloodstream, but these brief infections very rarely cause disease.
Diseases in which symptoms develop rapidly and resolve quickly (within a few days or weeks) are called acute infections. The common cold, for example, is an acute infection often caused by rhinoviruses and some coronaviruses. A chronic infection, however, is one in which symptoms develop gradually (over weeks or months) and are slow to resolve. Tuberculosis, a lung disease, is typically a chronic infection that can last for many months or even years if untreated.
How are pathogens classified regarding disease potential? Primary pathogens are disease-causing microbes that can breach the defenses of a healthy host. For example, the rod-shaped bacillus Shigella flexneri is a primary pathogen that causes bacillary dysentery. When ingested, it can survive the natural defensive barrier of an acidic stomach, enter the intestine, and begin to replicate. The organism in the chapter-opening case history, Treponema pallidum, is an example of a primary, or true, pathogen because it can survive initial attack by a healthy immune system and spread through the body in the bloodstream. Opportunistic pathogens, on the other hand, cause disease only in an immune-deficient host. The fungus Pneumocystis jirovecii (Figure 2.4A) easily infects people but rarely causes disease. It is, however, an opportunistic pathogen that causes life-threatening infections (opportunistic infections) in AIDS patients, whose immune systems have been eroded by HIV.
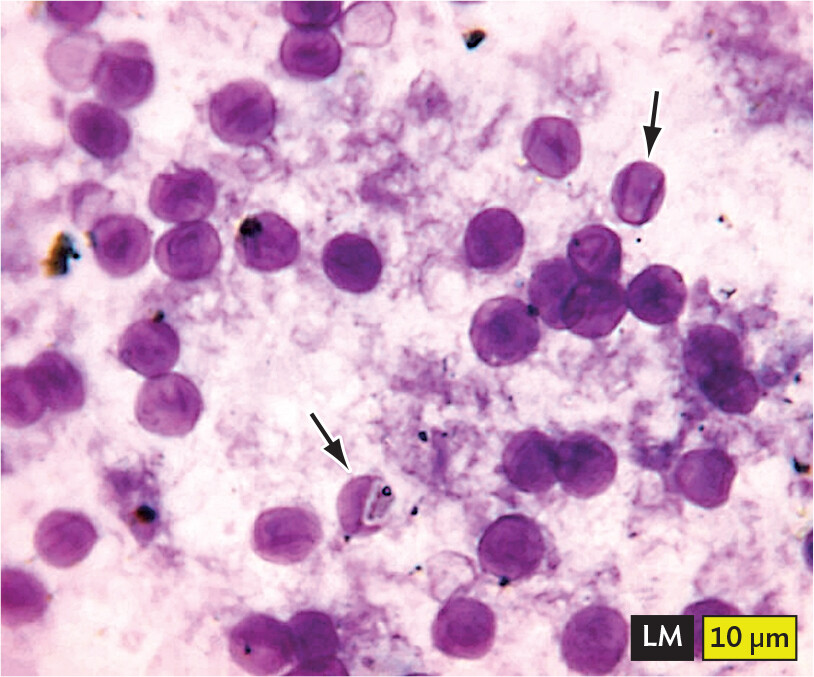
Figure 2.4Opportunistic and Latent Infections
More information
A light micrograph of Pneumocystis jirovecii cysts. The cysts are spherical, but have a thin, deflated quality that gives them an appearance similar to crushed Ping Pong balls. Darker material is visible within the cysts. The cysts are stained purple in this micrograph. Each cyst has a diameter of about 8 micrometers.
A.Pneumocystis jirovecii cysts (arrows) in bronchoalveolar material. Notice how the fungi look like crushed Ping-Pong balls.More information
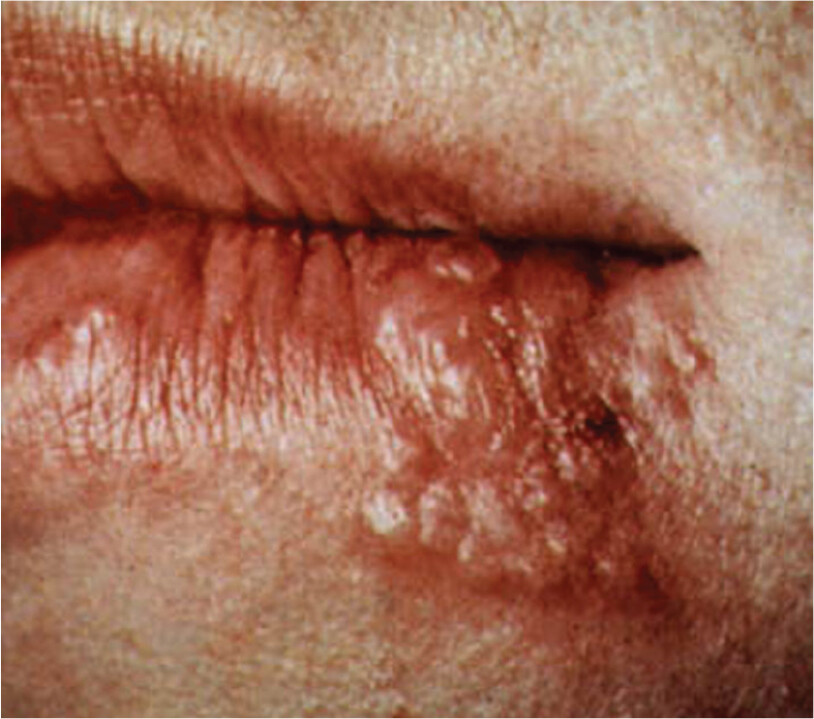
A photo of a herpes infection on the lips of a person. There is a cluster of tiny, fluid filled blisters on the lip and skin. The blisters and the surrounding skin are red and irritated.
B. Cold sore produced by a reactivated herpesvirus hiding latent in nerve cells.
Some microbes even enter into a latent state during infection, where the organism cannot be found by culture. Herpesvirus, for instance, can enter the peripheral nerves and remain dormant for years, then suddenly emerge to cause cold sores (Figure 2.4B). Another example, the bacterium Rickettsia prowazekii, causes epidemic typhus, but it can also enter a mysterious latent phase and emerge months or years later to cause a disease relapse called recrudescent typhus. Where in the body does this bacterium hide? With all our technology, evidence of a latent form of R. prowazekii has never been found—yet it must be there.
What distinguishes primary from opportunistic pathogens is their pathogenicity, defined by how easily the organism causes disease (its infectivity) and how severe that disease becomes (its virulence). Pathogenicity, overall, is shaped by the genetic makeup of the pathogen. In other words, an organism is more—or less—pathogenic depending on the tools at its disposal (such as toxins) and their effectiveness. Thus, the pathogenicity of a primary pathogen is greater than that of an opportunistic pathogen.
The virulence component of pathogenicity is a measure of the degree or severity of the disease. For instance, Ebola virus (Figure 2.5) and the closely related Marburg virus have case fatality rates of about 50%. This means that they are highly virulent. On the other hand, rhinovirus, the cause of the common cold, is very effective at causing disease but rarely kills its victims. So it is highly infective but has a low virulence. Both organisms are pathogenic, but with rhinovirus you probably live and with Ebola you probably die.
Figure 2.5 Highly Virulent Ebola Virus
More information
A micrograph of the Ebola virus and a photo of a healthcare worker attending to a patient with Ebola virus. The first part is the micrograph. The virus has a long and thin filamentous structure which forms loops and tangles. The second part is the photo. The health worker is wearing full P P E, including a face mask, face shield, gloves, and a full body protective suit. The patient is laying on a cot and appears to be experiencing discomfort.
A. Ebola virus.B. A health care worker tends to an Ebola patient in Kenema, Sierra Leone. Ebola causes hemorrhagic infections in which patients bleed from the mouth, nose, eyes, and other orifices. The mortality rate for this disease is approximately 50%.
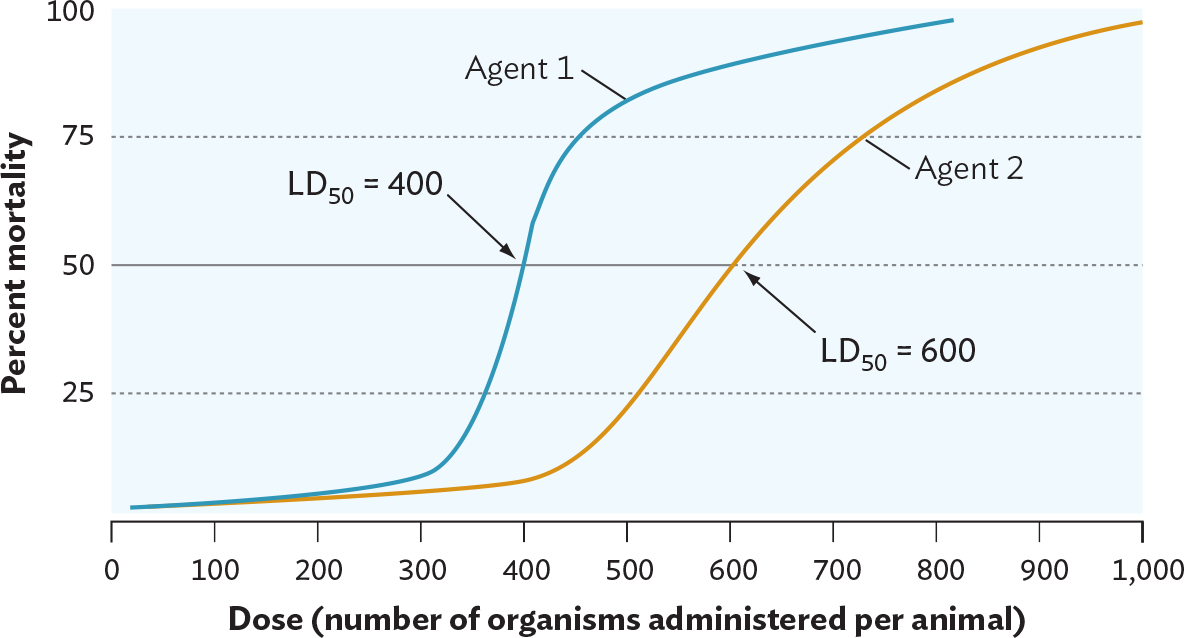
One way to measure virulence is to calculate a pathogen’s lethal dose 50% (LD50). The LD50 is the number of bacteria or virus particles (virions) required to kill 50% of an experimental group of animal hosts (usually mice or guinea pigs). An organism with a low LD50, in which very few organisms are required to kill 50% of the hosts, is more virulent than one with a high LD50 (Figure 2.6). For organisms that colonize but do not kill their hosts, a different measure called the infectious dose, or more properly the infectious dose 50% (ID50), is used. The ID50 is the number of organisms needed to colonize 50% of the experimental hosts. The ID50 is measured by determining how many microbes are required to cause disease symptoms in half of an experimental group of hosts.
Figure 2.6 Measurement of Virulence
More information
A line graph of the virulence of two different agents. The x axis displays dose in the number of organisms administered per animal from 0 to 1000. The y axis displays percent mortality from 0 to 100. Agent 1 reaches a high percent mortality at a lower dose than Agent 2. Agent 1 has an L D subscript 50 at a dose of 400. Agent 2 has an L D subscript 50 at a dose of 600. Both agents have a 0 percent mortality at a dose of 0. The percent mortality of Agent 1 slowly rises as the dose increases to 300. Between a dose of 300 and 400, the percent mortality of Agent 1 rapidly increases from 10 percent to 50 percent. Between a dose of 400 to 500, the percent mortality of Agent 1 continues to rise from 50 percent to about 80 percent. The percent mortality only increases slightly for doses higher than 500. The percent mortality of Agent 2 slowly rises as the dose increases from 0 to 450. Between a dose of 450 to 600, the percent mortality of Agent 2 steadily increases from about 10 percent to 50 percent. The percent mortality continues to gradually rise for doses higher than 600.
Each LD50 measurement requires infecting small groups of animals with increasing numbers of an infectious agent and observing how many animals die. The number of microbes that kill half the animals is called the LD50. In this example, agent 1, which requires fewer organisms to kill half the animals than does agent 2, is more virulent than agent 2.
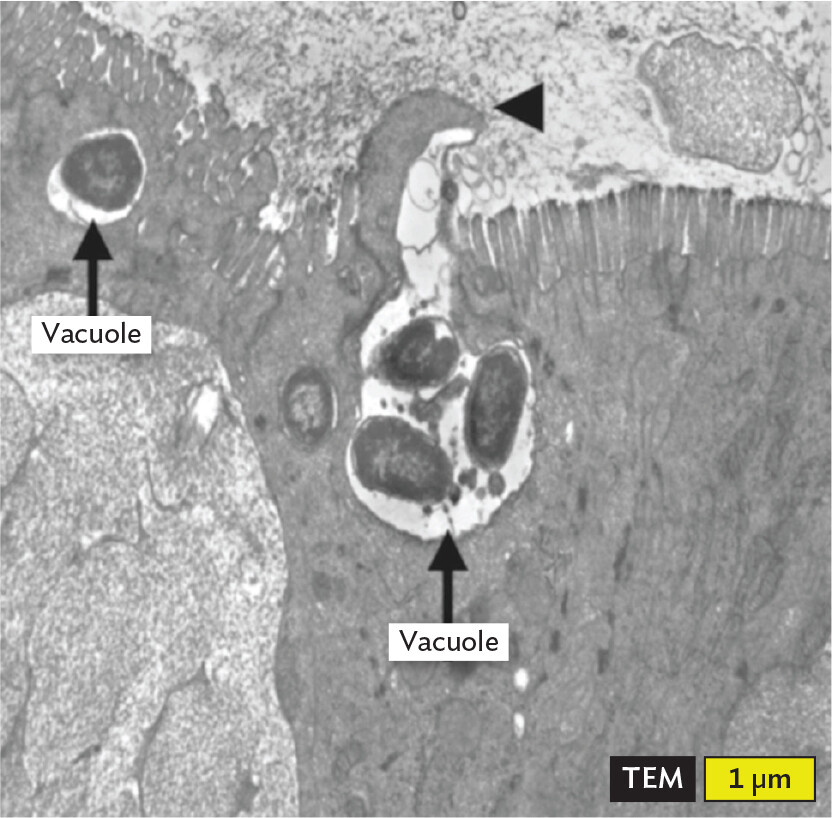
Two other aspects of pathogenesis are invasion and invasiveness. These terms sound similar but describe very different things. Invasion can mean the entry of a pathogen into a living human, or the process by which a single cell of a pathogen enters into (invades) an animal cell and grows there. Some pathogens live inside small, membrane-enclosed structures within the host cell called vacuoles (Figure 2.7A). Examples are Salmonella, a cause of diarrhea; Coxiella, the cause of Q fever; and Legionella, the cause of Legionnaires’ disease. Other bacteria prefer to live directly in the cytoplasm of the host cell, not in a vacuole (Figure 2.7B). Examples of bacteria that grow in cytoplasm are Shigella, a cause of bloody diarrhea, and Listeria, which can cause diarrhea and a lethal neonatal meningitis.
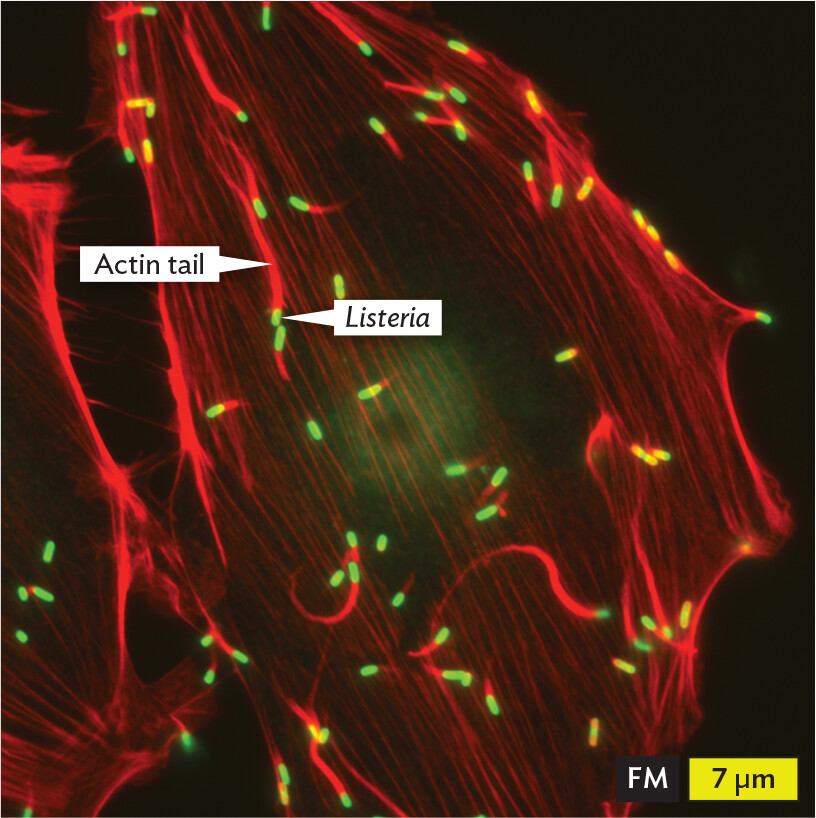
Figure 2.7Bacterial Life in a Host Cell
More information
A transmission electron micrograph of Salmonella enterica bacterial cells invading intestinal cells. The micrograph is in greyscale. The field of view is centered on vacuoles inside of an intestinal cell. Several spherical to ovoid cells are seen in the vacuoles. The cells have dark grainy interiors. Each cell is around 1 micrometer in maximum dimension.
A. Invasion of intestinal cells (enterocytes) by Salmonella enterica serovar Typhimurium. Intracellular bacteria are located within membrane-enclosed vacuoles (arrows). Bacteria are first captured by cytoplasmic projections coming from the enterocyte surface (arrowhead).More information
A fluorescence micrograph of Listeria bacterial cells invading eukaryotic host cells. The Listeria cells fluoresce green. The cells are rod shaped and have long actin tails extending behind them. The actin tails fluoresce red. The cells travel in various directions across the field of view, leaving long red trails across a dark background. Each Listeria cell is about 1 micrometer in length and 0.5 micrometer in width.
B. The bacterium Listeria similarly invades eukaryotic host cells by being engulfed in a vacuole, but the pathogen then escapes the vacuole to live in the cytoplasm. Listeria (green) can propel itself in the cytoplasm by linking together (polymerizing) many copies of a host protein (actin) at one end of the bacterium. The structure is called an actin tail (red). Regions where the red and green fluorescence overlap appear yellow. The host cell is invisible in this image but encompasses the bacteria.
Invasiveness, in contrast to invasion, refers to the ability of a bacterial pathogen to rapidly spread through tissue without invading host cells. For instance, some strains of Streptococcus pyogenes are said to be highly invasive because they secrete enzymes that degrade host tissues. These strains are sometimes called “flesh-eating bacteria” because of how quickly they destroy tissue as the organism spreads.
Note: The disease processes and pathogenic mechanisms of the many microbes mentioned in this chapter will be explained more completely in later chapters. The basic concepts described here, however, can be applied to all the case histories that anchor the chapters of this book.
Different microbes also have different host ranges that describe which animals they can infect and produce disease in. Pathogens can have a very narrow host range; for example, Salmonella enterica serovar Typhi, the bacterium that causes typhoid fever, can infect only humans. (Serovar is a subclass within a species.) But a close relative, Salmonella enterica serovar Typhimurium, which produces diarrhea in humans, can infect many different animals. S. Typhimurium, then, has a broad host range. T. pallidum, from the chapter-opening syphilis case, has a very narrow host range. It infects only humans.
Note: Bacterial names include designations for genus (Salmonella) and species (enterica). Both are written in italics, with the genus capitalized and species lowercase. Serovar subclass names of a species are not italicized but are capitalized (for instance, Typhimurium).
When Pathogen Meets Host
All pathogens—whether viral, bacterial, or eukaryotic—must interact with a host in several ways to be successful. The pathogen must attach to a host tissue, avoid the host’s immune system, and steal nutrients from the host. We will delve more deeply into these interactions later (especially in Chapters 18–24), but let’s discuss the basic concepts here.
Attachment. Specific proteins (adhesins) on the surface of microbes help them adhere to host cells, like thumbtacks to a corkboard. Microbial adhesins bind to host cell surface structures called receptors (proteins, lipids, or polysaccharides) that have roles critical to normal host physiology. Because different host cells in different tissues have different cell-surface structures (in other words, different receptors), a pathogen will preferentially infect tissues whose cells are “decorated” with the right surface receptors. Pathogens that fail to attach to the host can be expelled from the body by coughing, urinating, or defecating.
Immune avoidance. Different pathogens possess unique tools to avoid the immune system. Some, such as Salmonella, periodically change the molecular shape of their surfaces to confuse the immune system. Others, such as Staphylococcus aureus, secrete molecules that “tell” immune system cells, “All is well; no infection here.” Still others (Shigella, for instance) secrete proteins that convince immune system cells to kill themselves in a process known as apoptosis (pronounced “ay-pop-TOH-sis”). Several of the ingenious strategies that microbes use to evade the immune system will be described in Chapters 15 and 16.
Stealing nutrients from the host. Animals, including humans, have evolved ways to safeguard nutrients that bacteria need in order to grow. One example involves iron. To keep iron away from invading bacteria, the human body makes compounds (lactoferrin and transferrin) that tightly bind this metal. Consequently, most bacteria that enter the bloodstream cannot access the iron in the blood and cannot grow. Pathogens, however, have ways of stealing the iron from host tissues to use for their own purposes (see Section 5.2).
SECTION SUMMARY
Infection with a microbe does not always lead to disease.
Primary pathogens have mechanisms that help the organism circumvent host defenses.
Opportunistic pathogens cause disease only in a compromised host.
Pathogenicity refers to a pathogen’s disease potential, defined by its ease of entering a host (infectivity) and disease severity (virulence).
Invasion refers to the entry of a pathogen into host cells, where it then lives. Invasiveness describes the ability of an organism to spread through tissues.
Host range refers to how readily a pathogen can infect different animals.
Pathogens must be able to attach to host cells, evade the immune system (at least temporarily), and obtain nutrients from the host.
Thought Question 2.1 Is a microbe with an LD50 of 5 × 104 more or less virulent than a microbe with an LD50 of 5 × 107? Why?
SHOW ANSWERHIDE ANSWER
Because it takes fewer cells to cause disease, the microbe with the smaller LD50 (5 × 104 ) is the more virulent one.
Thought Question 2.2 The bacterium Salmonella enterica serovar Typhimurium (S. Typhimurium) is usually transmitted to humans by contaminated food and causes a diarrhea that typically resolves without medical intervention. During infection, the organism remains localized to the intestine. A different serovar of Salmonella, S. Typhi, is also transmitted via contaminated food but can penetrate the intestinal wall and enter the lymphatic system and bloodstream, causing high fever and, in some cases, death. Clearly, S. Typhi is more virulent than S. Typhimurium to humans. However, when tested for lethal dose (LD50) in mice, S. Typhi fails to kill any mice, whereas S. Typhimurium has a very low lethal dose (it takes very few organisms to kill mice). How might you broadly explain this apparent contradiction?
SHOW ANSWERHIDE ANSWER
S. Typhi is host adapted to infect only humans. It will not infect other hosts, such as mice, because the bacterium either cannot attach to host cells in other animals (the right receptors are missing) or is unable to counter their immune mechanisms. S. Typhimurium has a broader host range but lacks some of the tools that S. Typhi uses to cause severe disease in humans.
A bacterial, viral, fungal, protozoan, or helminthic agent of disease; among health professionals, pathogens typically are limited to bacteria, viruses, and fungi.
Any bacterium, virus, fungus, protozoan (protist), or helminth that colonizes and harms its host; the term commonly refers to protozoa and to invertebrates.
 Note: The various types of microscopy used to see microbes will be described in
Note: The various types of microscopy used to see microbes will be described in