This part of the chapter considers vaccination, a routine medical procedure that gives people an immunological memory of a pathogen without having to be infected and suffer the disease it causes. This ‘miracle’ is achieved by immunizing people with noninfectious material, called the vaccine, which contains the pathogen’s antigens. Two or more immunizations are usually given. The first mimics a first infection by the pathogen and stimulates a primary immune response. Given at a later time, the second immunization stimulates a secondary immune response to the vaccine, and so on for additional immunizations. Because of the immunological memory induced by vaccination, any subsequent infections with the pathogen are swiftly eliminated before they cause disease. To protect individuals and populations from infectious disease, children should be vaccinated when they are young. Vaccination has protected billions of people from infection and is by far the most successful and frequently used manipulation of the immune response. Vaccination has probably saved more human lives than any other medical procedure.
The modern era of vaccination began in 1796 with Edward Jenner’s report on using material from animals infected with cowpox, a disease of cattle, as a vaccine to protect against smallpox. Although Jenner’s procedure was widely embraced, vaccines for other diseases did not emerge until well into the 19th century—the era of the microbe hunters, when microorganisms were first isolated, grown in culture, and shown to cause disease. Mainly by trial and error, methods were developed to inactivate pathogens, or impair their capacity to cause disease, and these crippled microbes were used as vaccines. Eventually, vaccines were developed against most of the epidemic infections that then plagued humankind in western Europe and North America. With the discovery of antibiotic drugs in the mid-20th century, it seemed that the combination of vaccines and antibiotics would solve for good the problems posed by infectious disease. Such optimism was soon tempered by the difficulties encountered in developing effective vaccines for some diseases and by the emergence of new pathogens and of antibiotic-resistant strains of the old ones. The sequencing of pathogen genomes and the application of recombinant DNA technology have, however, opened up new and exciting prospects for making vaccines.
11-11Protection against smallpox is achieved by immunization with the less dangerous vaccinia virus
The first medically prescribed vaccine was against smallpox, a viral disease characterized by a rash of spots that become virus-loaded pustules, leaving permanent scars on the skin of survivors. The rash frequently involved the face, because the disease was mainly spread by face-to-face contact. The earliest vaccines actually contained pathogenic smallpox virus; they were made from dried pustules taken by physicians from patients experiencing a mild form of the disease. Small amounts of this material were given to healthy people, including children, either intranasally or intradermally through a scratch on the arm—a procedure known as variolation, from the word variola, the Latin name given both to the pustule and to the disease itself. Although successful in many cases, the drawback of variolation was the frequency with which it produced full-blown smallpox, resulting in the death of around 1 in 100 of those vaccinated. Despite the risk, variolation was widely used in the 18th century because the threat from smallpox was so much greater than that from variolation. At the time, smallpox killed one in four of those infected, and epidemics occurred on a regular basis. In London, for example, more than one-tenth of all deaths were due to smallpox.
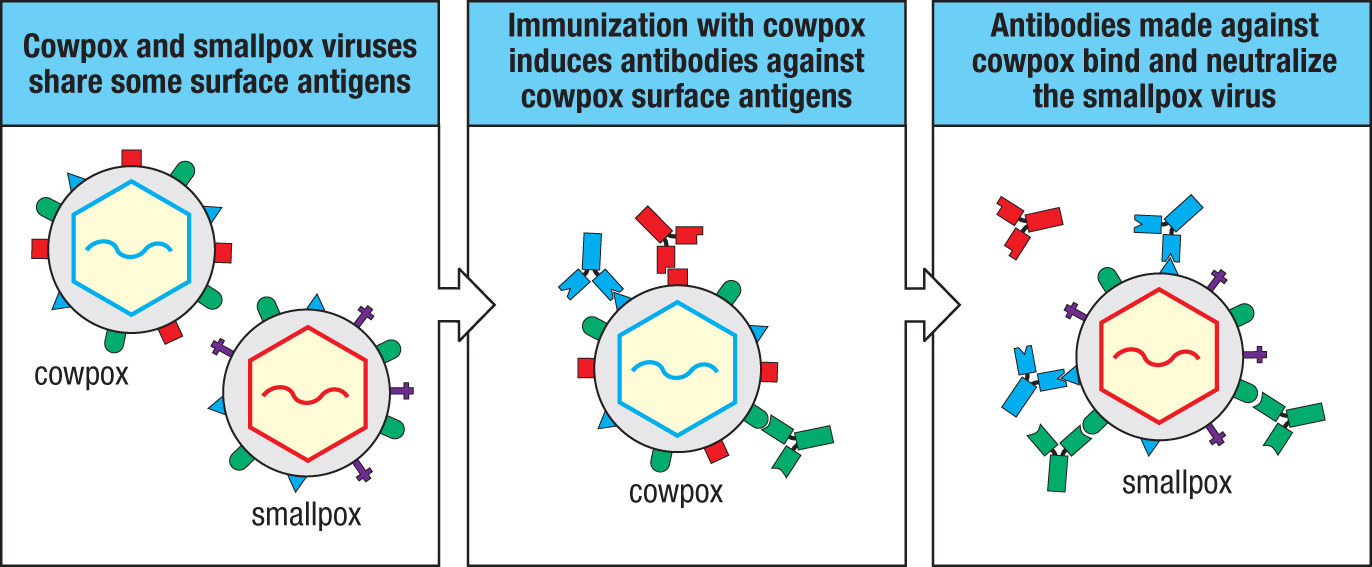
Toward the end of the 18th century, Jenner’s innovation was to use the related cowpox virus as a vaccine for smallpox. The cowpox virus causes very mild infections in people, but the immunity produced gives effective protection against both smallpox and cowpox because the two viruses have antigens in common (Figure 11.16). In the 19th century, Jenner’s vaccine replaced variolation and more modern vaccines using the related vaccinia virus were eventually responsible for the eradication of smallpox in the 20th century. The words ‘vaccinia’ and ‘vaccination’ derive from vaccus, the Latin word for ‘cow.’ Although the term vaccination was originally used in the context of smallpox, it now refers to any immunization that induces protective immunity against a disease.
Figure 11.16Vaccination with cowpox virus elicits neutralizing antibodies against epitopes shared with smallpox virus. Cowpox also elicits T-cell immunity that cross-reacts with smallpox (not shown).
11-12Smallpox is the only infectious disease of humans that has been eradicated worldwide by vaccination
Immunization with vaccinia induces long-lasting immunity (see Figure 11.1) and proved extremely effective against smallpox, leading to the worldwide eradication of the disease in 1979. After that, the immunization of children against smallpox began to be discontinued, and a smallpox vaccination is no longer part of the recommended childhood vaccinations in the United States (Figure 11.17). Smallpox is the only epidemic infectious disease to have been globally eradicated by vaccination, which was achieved through a coordinated international program. Three properties of the smallpox virus contributed to this success. As smallpox evolves slowly and has conserved antigenic epitopes, the immunity acquired as an infant remains effective throughout life. The vaccine is a live virus that establishes an infection at the injection site in the skin, the tissue where natural smallpox infections occur. Thus the innate and adaptive immune responses stimulated by the vaccine mimics smallpox infection and produces immunological memory that is effective against smallpox. Last, but not least, as smallpox only infects the human species, there are no reservoirs of the virus in other animals. Once the chain of transmission among humans was broken, the virus could not survive.
Figure 11.17Childhood vaccination schedules in the United States. Each red box denotes a time at which a vaccine dose should be given. Boxes spanning multiple months indicate a range of times during which the vaccine may be given. DTaP, diphtheria, tetanus, and acellular pertussis vaccine. Based on Centers for Disease Control and Prevention immunization schedules.
Current immunization schedule for children (USA)
Vaccine given
Birth
2 months
4 months
6 months
12 months
15 months
18 months
4-6 years
11-12 years
14-16 years
Hepatitis B
Rotavirus
Diphtheria–tetanus–pertussis (DTaP)
Haemophilus B conjugate (HiBC)
Pneumococcal conjugate (PCV13)
Inactivated polio vaccine
Influenza
Annual vaccination
Measles/mumps/rubella (MMR)
Varicella (chickenpox virus)
Meningococcus C
Human papillomavirus
An added benefit of smallpox vaccination was the protection it gave against other poxviruses, such as cowpox and monkeypox, which do infect humans and cause mild disease. In the absence of routine smallpox vaccination, human immunity to the other poxviruses began steadily to wane, while the frequency of human infections with these other poxviruses, such as monkeypox in West Africa, began steadily to increase. From 1978 to 2016 there were no reported cases of monkeypox in Nigeria. An outbreak of monkeypox occurred in 2017 in which 116 people were infected and 8 of them died. Since then, the number of cases has risen exponentially and people in 10 African countries have been affected.
11-13Most viral vaccines are made from killed or inactivated viruses
Jenner’s strategy for smallpox vaccination has not been applicable to most pathogenic viruses because they lack a ‘safe’ natural relative. Most viral vaccines contain the disease-causing virus, but in a form that does not cause severe disease. One type of vaccine consists of virus particles (virions) that cannot replicate because they are treated chemically with formalin or physically with heat or irradiation. These are called killed (or inactivated) virus vaccines. The influenza and rabies vaccines are of this type. The strategy only works if the viral nucleic acid can be completely and reliably inactivated. This is not trivial because of the large quantities of pathogenic virus produced during vaccine manufacture.
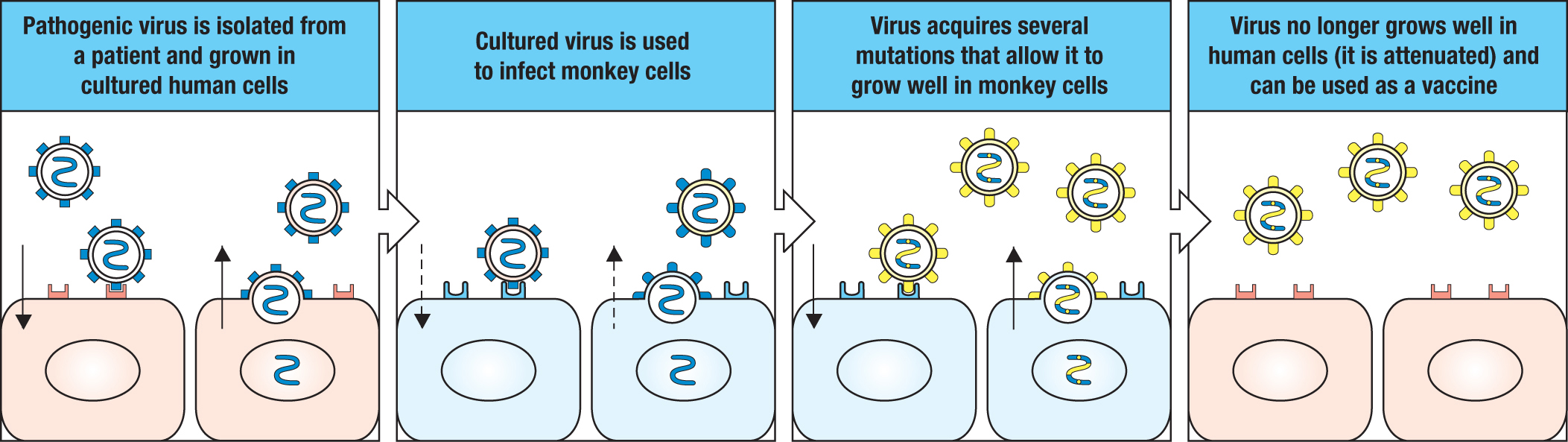
A second type of vaccine comprises a mutant form of live virus that grows poorly in human cells and is no longer pathogenic to humans. Such vaccines, called live-attenuated virus vaccines, usually elicit a better protective immunity than that by killed virus vaccines. This is because the attenuated virus has a limited capacity to infect cells and replicate, and thereby mimics a real infection. Most of the viral vaccines currently used to protect humans are live-attenuated vaccines. To attenuate a pathogenic human virus, it is grown in cells from another animal species. Such conditions select for variant viruses that grow faster in the non-human cells, which makes them less fit for growth in human cells (Figure 11.18). The measles, mumps, and yellow fever vaccines contain live-attenuated viruses. Attenuated viral strains can also arise naturally. As polio infection passes though a human population, the poliovirus diversifies by mutation, occasionally producing a strain with reduced pathogenicity. Such natural live-attenuated viruses are suitable candidates for vaccines, and one such strain of poliovirus (strain 2) is a part of the oral polio vaccine.
Figure 11.18Attenuated viruses are obtained by growing pathogenic human viruses in non-human cells. To produce an attenuated form of a virus, the virus is first grown in quantity in human tissue culture cells. The virus is then isolated and grown in cells of non-human species, such as monkey species, until it adapts fully to the non-human cells and as a consequence grows poorly in human cells. The adaptation is the result of selection for a mutant virus that outgrows the original virus. The attenuated mutant usually differs from the original virus by an accumulation of several point mutations. Although growing slowly in the human host, the attenuated virus of a vaccine is present in amounts that stimulate an immune response but are insufficient to cause disease. The rubella vaccine is the one vaccine that was produced by attenuation during culture in human cells.
11-14Both inactivated and live-attenuated vaccines protect against poliovirus
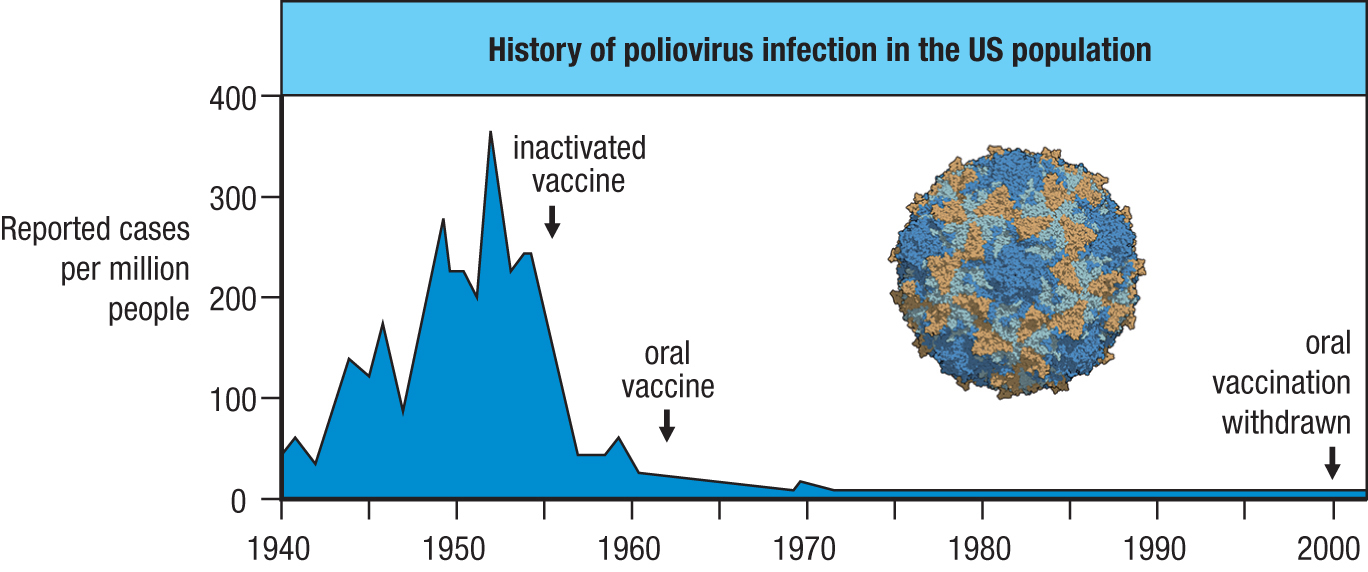
Although less than 1% of infections with poliovirus cause disease, this minority produces a variable paralysis called poliomyelitis, or infantile paralysis, that can kill or disable the affected individuals, most of whom are children. Polio was a major epidemic disease in the 20th century and was particularly common in the United States, where the worst epidemic on record occurred in 1952. Whereas smallpox is transmitted by face-to-face contact, poliovirus is transmitted by the fecal–oral route, leading to various disease outbreaks being associated with communal bathing in public swimming pools. Because the virus infects gut mucosal surfaces, this indicated that an oral vaccine would be superior to an injected vaccine. But the challenge for any oral vaccine is to prevent its degradation in the gut before it reaches its target, the intestinal epithelium. The first polio vaccine (the Salk vaccine) was approved in 1955; it contained killed virus of the three main strains and was injected into the skin. Because it contained three different polioviruses, this vaccine was said to be trivalent. By 1963, a similarly trivalent oral live-attenuated virus vaccine (the Sabin vaccine) was introduced. Because this trivalent oral polio vaccine (TVOP) gave better protection and was easier to administer, it began to replace the killed virus vaccine. Together, the two vaccines achieved an impressive reduction in the incidence of polio in the United States, where the last recorded case was in 1979 (Figure 11.19). The Global Polio Eradication Initiative began in 1988, when polio was endemic in 125 countries and caused the paralysis of 350,000 children each year. By the year 2000, the number of cases of polio was reduced by 99%, and since then there has been further decrease. Poliovirus has been eliminated from all countries except Pakistan and Afghanistan. Because poliovirus persists in those countries and is highly contagious, children in other countries are still being vaccinated against polio (see Figure 11.19). As poliovirus, like smallpox, infects only humans, there is potential for its complete elimination from the human species. If that goal is reached, poliovirus will then be extinct in the ‘wild’ and restricted to a few research laboratories.
Figure 11.19A successful vaccination campaign. Poliomyelitis has been virtually eliminated from the United States, as shown by the graph. The first arrow shows when the campaign to eradicate polio began in the 1950s with introduction of the inactivated vaccine. The second arrow marks the introduction of the attenuated oral vaccine in the 1960s. In the last 30 years of the 20th century, polio had become a rare disease in which public concern turned away from epidemic polio and toward the cases of polio caused by the attenuated viral strains of the oral vaccine. The third arrow shows when the oral vaccine was withdrawn in the year 2000. Because poliovirus has not been eradicated worldwide and the volume of international travel is so high, immunization must be maintained in a good majority of the US population in order to prevent recurrence of epidemic disease. The poliovirus capsid is shown.
11-15Vaccination can inadvertently cause disease
Soon after the Salk polio vaccine was introduced in 1955, an outbreak of polio occurred among people who had received the vaccine and their families and friends. This disaster immediately caused polio vaccination in the United States to be suspended for several months while its cause was investigated. All the infections were traced back to one manufacturer and two batches of vaccine containing pathogenic virus that was not completely inactivated. Some 200,000 people received the live virus. Of these 70,000 became ill, 200 were permanently paralyzed, and 10 died. Much of the public’s current distrust of vaccination stems from this calamity.
After its introduction and extensive use, the Sabin vaccine was found to induce polio and paralysis in three people per million vaccinated. These infections are all caused by strain 3, one of three live-attenuated polio strains that make up TVOP. All three of the strains are necessary to provide adequate protection against the natural range of poliovirus variants. Considering just the numbers, the inclusion of strain 3 in the vaccine prevents many more cases of disease than it causes. Although the attenuated strain 3 virus differs from a natural pathogenic strain of poliovirus by 10 nucleotide substitutions, a back-mutation at just one of these positions is sufficient for strain 3 to revert and become pathogenic. Such a mutation in the viral genome can occur during either vaccine manufacture, when it should be caught and eliminated during quality control, or after vaccination, when the virus replicates in vaccinated people and causes disease.
In countries where poliovirus has been eradicated and the only cases of disease are caused by vaccination, there can be greater fear of the vaccination than of the disease. For this reason there has been considerable social pressure to improve the polio vaccine, and some parents and communities refuse to have their children vaccinated. One way to reduce the exposure to strain 3 is to use an inactivated polio vaccine (IPV) for the first immunization, and to follow this with TVOP for the second and subsequent immunizations. In this way the primary immunization cannot lead to infection, and strain 3–specific antibodies made in the primary response help to prevent infection by any revertant of strain 3 during the subsequent immunizations. Because of the reversion rate, in the year 2000 TVOP was no longer recommended for routine mass vaccination in the United States, and IPV became the vaccine of choice (see Figure 11.19).
In 2007, the people of northern Nigeria experienced one of the largest recorded outbreaks of polio caused by a vaccine strain. Over a 2-year period, 69 children were paralyzed as a result of vaccination. In the two countries where polio remains endemic, episodes such as this have exacerbated the public’s fear and distrust of vaccination and also of the people administering the vaccine. Exploiting the situation are political extremists, who shot dead some 30 polio workers in Pakistan and Nigeria during 2013. Despite these setbacks, in 2018 there were only 8 cases of polio in Pakistan compared to 39 in 2015. Although Syria had been free of polio since 1999, social disruption caused by civil war led to reduced vaccination rates and an outbreak of 10 cases of polio in 2013 among infants less than 2 years old. In 2018, an outbreak of polio occurred in the Democratic Republic of the Congo (DRC), a country where wild-type polio is extinct. This widespread outbreak was caused by a recently evolved mutant of the vaccine strain that is as pathogenic as the wild-type strain. Complicating medical efforts to respond to the polio outbreak was an outbreak of Ebola virus disease in the DRC that began in August 2018. By May 2019, Ebola had been diagnosed in 1510 patients and 994 of them had died. In such volatile and difficult environments polio can flourish, reducing the likelihood that it will soon be globally eradicated.
11-16Subunit vaccines are made from the most antigenic components of a pathogen
In the immune response to hepatitis B virus (HBV) the protective, neutralizing antibodies are made against the surface protein of the virus, also called the HBV surface antigen. The hepatocytes of patients infected with HBV secrete the surface protein into the blood as minute particles. The first version of the anti-hepatitis B vaccine contained surface protein that was purified from the plasma of HBV-infected people. Because only one component, or subunit, of the virus is used for vaccination, this type of vaccine is called a subunit vaccine. In purifying the antigenic particles, the greatest concern was that infectious particles of HBV should be completely removed. If they were not, vaccination could infect people with the virus and give them disease, instead of protecting them from it.
This concern, and the proven benefit of the vaccine in preventing life-threatening liver disease, led to the HBV subunit vaccine being one of the first vaccines to be made using recombinant DNA technology. Making the vaccine in this way did not involve viral particles, and it avoided any contact with them. The gene encoding the HBV surface antigen was inserted into the genome of baker’s yeast. The recombinant yeast was then grown in mass cultures, from which the surface protein was purified in large quantity. The vaccine was introduced in 1986 and provides protective immunity against infection by HBV to about 85% of the people vaccinated.
11-17Invention and application of rotavirus vaccines took decades of research and development
The wheel-shaped rotavirus was discovered in 1973 and shown to be a major cause of severe childhood diarrhea. Almost all children are exposed to rotavirus, and in 2008 the illness it caused was responsible for more than 2 million stays in hospitals and around 450,000 deaths per year in children under 5 years of age worldwide. Although efforts to make a vaccine began soon after the virus was discovered, it took more than 30 years to develop a vaccine that is effective and for which the side effects are acceptable. Two vaccines, Rotarix and RotaTeq, are currently used. These vaccines have resulted in a considerable decrease in mortality (Figure 11.20).
nd, not determinedFigure 11.20Childhood mortality from rotavirus infection in the 10 most affected countries, 2008 and 2016. The RotaTeq and Rotarix vaccines were licensed in 2006 and 2008, respectively. By 2017, it was estimated that 28% of 1-year-old children worldwide were vaccinated against rotavirus. Shown here for years 2008 and 2016 are the estimated annual numbers of deaths from rotavirus infection in children under 5 years old for the 10 countries having the highest mortality in those years. The ranking of the top five countries did not change from 2008 to 2016, but there has been an impressive and widespread reduction in mortality. Also shown for each country is the percentage of 1-year-old children who have been vaccinated. ‘nd’ indicates no data available, usually because rotavirus vaccination was not introduced by 2017. Empty boxes indicate countries that were not in the top 10 in either 2008 or 2016. Source of mortality data: 2008, World Health Organization (WHO); 2016, C. Troeger et al. 2018. JAMA Pediatrics 172: 958–965. Source of vaccination data: WHO, https://apps.who.int/gho/data/node.main.ROTACn?lang=en.
Annual childhood mortality from rotavirus infection the top 10 countries in 2008 and in 2016
Country
Annual deaths
Vaccine coverage of 1-year-olds (%)
2008
2016
2017
India
99,000
10,000
13
Nigeria
41,000
48,000
nd
Pakistan
39,000
2,000
12
Democratic Republic of the Congo
33,000
12,000
nd
Ethiopia
28,000
3,000
80
Afghanistan
25,000
nd
Uganda
11,000
2,000
nd
Bangladesh
10,000
nd
Indonesia
10,000
2,000
nd
Angola
9000
2,000
58
Chad
4000
nd
Niger
4000
84
Kenya
1500
67
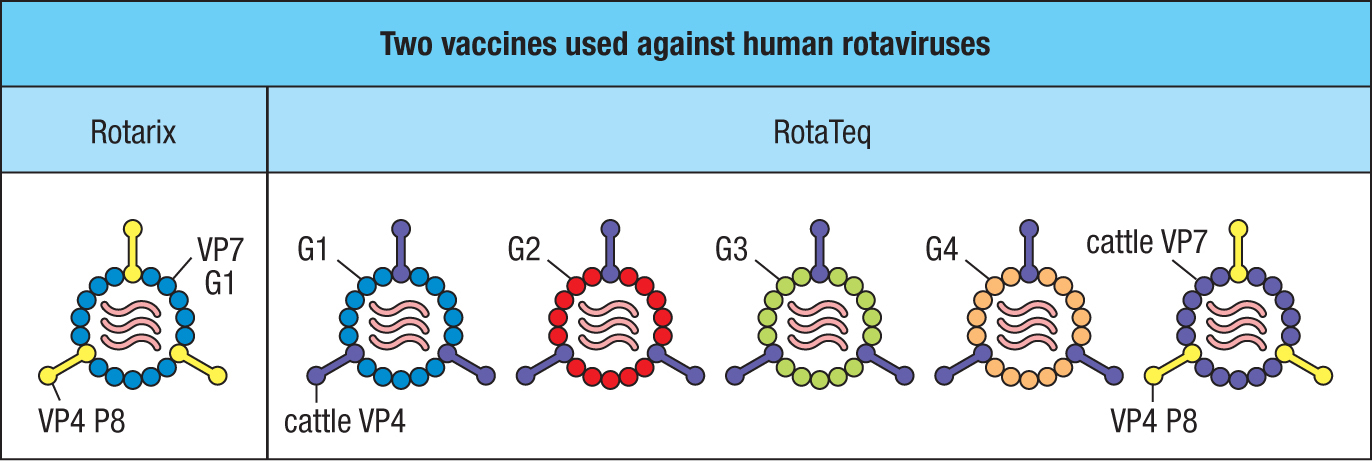
The rotavirus genome consists of 11 double-stranded RNA molecules. The VP4 and VP7 viral coat proteins are the main targets for neutralizing antibodies. Rotaviruses frequently mutate, with VP4 and VP7 being variable proteins that define different viral serotypes. The serotypes are further diversified by reassortment of the 11 genomic segments. Although there are 42 natural variants of rotavirus, five account for 90% of disease, and their antigens are incorporated into both vaccines. Rotarix is an attenuated human rotavirus containing VP4 and VP7. RotaTeq vaccine contains five cattle rotaviruses, each engineered to express a different human VP4 or VP7 variant (Figure 11.21). These vaccines give 85–98% protection against severe rotavirus diarrhea. Like the oral poliovirus vaccine, they can be made using simple tissue-culture techniques and are readily transferable to the less-developed African and Asian countries, where most mortality from rotavirus infection occurs (see Figure 11.20).
Figure 11.21Two effective rotavirus vaccines. The Rotarix vaccine (left panel) is an attenuated human rotavirus that has common P8 and G1 variants of the VP4 and VP7 glycoproteins, respectively. The RotaTeq vaccine is based on a cattle rotavirus that is nonpathogenic to humans. The vaccine consists of five viral strains, four expressing different human VP7 variants (G1, G2, G3, and G4) and cattle VP4, and one expressing cattle VP7 and the human P8 variant of VP4. Cattle components are in purple.
11-18Bacterial vaccines are made from whole bacteria, secreted toxins, or capsular polysaccharides
The development of vaccines against bacterial pathogens used similar strategies to those outlined in the previous sections for viral pathogens. The first live-attenuated bacterial vaccine to prove effective was derived from a bovine strain of Mycobacterium tuberculosis and is effective against tuberculosis (TB). This Bacille Calmette–Guérin (BCG) strain is not pathogenic to healthy humans. The vaccine’s efficacy varies with the human population, and although routinely given to children in some European countries, BCG has never been used in the United States. More recently, live-attenuated vaccines against species of Salmonella have been introduced for both medical and veterinary use. An attenuated strain of Salmonella enterica serovar Typhi, the cause of typhoid fever, was made by mutagenesis and selection for the loss of a lipopolysaccharide necessary for pathogenesis. In this vaccine strain, an enzyme necessary for lipopolysaccharide synthesis is defective.
Some bacterial diseases are caused entirely by toxic proteins secreted by the bacteria. Most important of these are the diphtheria toxin of Corynebacterium diphtheriae that causes diphtheria and the tetanus toxin of Clostridium tetani that causes tetanus. To avoid contracting these diseases, infected individuals need high-affinity neutralizing antibodies that bind irreversibly to the toxin and inhibit its toxic activity. In the presence of such antibodies, the toxin molecules are neutralized the moment they are secreted, before they can affect human cells and cause disease (see Section 9-16). Such vaccines are made by treating purified toxins with formalin to destroy their toxicity. The inactivated proteins, called toxoids, retain sufficient antigenic activity to stimulate the production of neutralizing antibodies. Diphtheria and tetanus vaccines are thus comparable to the viral subunit vaccines.
Many pathogenic bacteria have an outer capsule composed of polysaccharides that define species-specific and strain-specific antigens. For these encapsulated bacteria, which include the pneumococcus (Streptococcus pneumoniae), salmonellae, the meningococcus (Neisseria meningitidis), Haemophilus influenzae, Escherichia coli, Klebsiella pneumoniae, and Bacteroides fragilis, the capsule determines both the pathogenicity and the antigenicity of the organism. In particular, the capsule prevents the fixation of complement by the alternative pathway. Only when antibodies have bound to the capsule does complement fixation lead to bacterial clearance. Consequently, the aim of vaccination against such bacteria is to produce complement-fixing antibodies that bind to components of the capsule.
11-19Conjugate vaccines enable high-affinity antibodies to be made against carbohydrate antigens
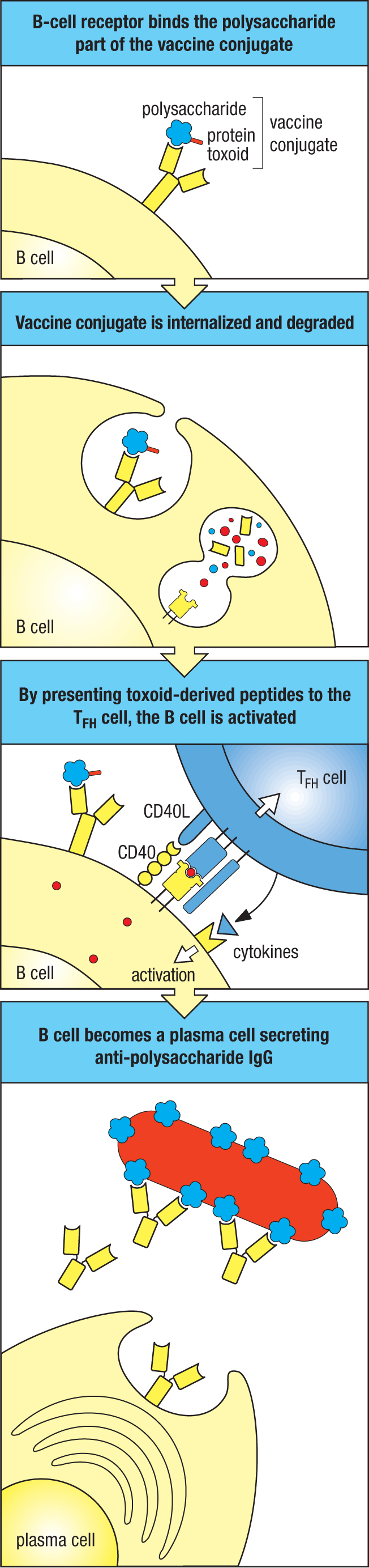
During the 1990s, an epidemic of meningitis in Europe and North America was due to a hypervirulent form of serogroup C N. meningitidis. This bacterium colonizes most people with no harmful effect, but it invades the blood and the brain of some individuals, causing disease that can either be fatal or cause permanent damage to organs and tissues. The disease mainly affects young children of around 2 years of age and adolescents of 15–19 years. Protection against this infection is provided by high-affinity IgG that binds to polysaccharides of the capsule and triggers complement-mediated killing of the bacteria. At the time of the epidemic, the existing vaccine proved ineffective at protecting infants. The vaccine’s deficiency was that it contained bacterial polysaccharides but no bacterial proteins. Consequently the vaccine failed to activate helper CD4 TFH cells and could only stimulate a T-independent B-cell response that gave rise to low-affinity IgM, with no potential for somatic hypermutation, isotype switching, or memory B cells (see Section 9-3). It was failures such as this that led to the concept of conjugate vaccines, in which different epitopes recognized by B cells and T cells are synthetically linked together (Figure 11.22).
Figure 11.22Macromolecular complexes with separate epitopes recognized by B cells and T cells make effective vaccines. The first panel shows a naive B cell’s surface immunoglobulin binding a carbohydrate epitope on a vaccine composed of a Haemophilus polysaccharide (blue) conjugated to tetanus toxoid (red), a protein. This results in receptor-mediated endocytosis of the conjugate and its degradation in the endosomes and lysosomes, as shown in the second panel. Peptides derived from degradation of the tetanus toxoid part of the conjugate are bound by MHC class II and presented on the B-cell surface. In the third panel, the receptor of a TFH cell recognizes the peptide:MHC complex. This induces the T cell to secrete cytokines that activate the B cell to differentiate into plasma cells, which produce protective antibody against the Haemophilus polysaccharide (fourth panel).
Effective N. meningitidis vaccines were made by conjugating the bacterial polysaccharide to either tetanus or diphtheria toxoid. These inactive forms of toxic bacterial proteins were already being used in the tetanus and diphtheria vaccines and were known to stimulate a potent and neutralizing IgG response. On immunization, dendritic cells process the vaccine and present toxoid-derived peptides to naive CD4 T cells, which activate and differentiate into TFH cells. Naive B cells that are specific for epitopes of the bacterial polysaccharide endocytose the intact conjugate, and after processing the toxoid they present its peptide antigens to the activated toxoid-specific TFH cells. This interaction drives B-cell activation, leading to a germinal center reaction and the production of high-affinity, neutralizing IgG antibodies against the bacterial polysaccharide and the development of immunological memory (see Figure 11.22). At the time, 1998–1999, when vaccination with the conjugate began in the United Kingdom, there were 411 diagnosed infections with hypervirulent serogroup C N. meningitidis and 32 deaths of children aged 10 years or younger. In the ninth year of the vaccination program (2007–2008), only four infections occurred and no deaths. Similar success was obtained with conjugate vaccines against two other pathogenic species of encapsulated bacteria: H. influenzae and S. pneumoniae.
11-20Adjuvants are added to vaccines to activate and enhance the immune response to a pathogen
Vaccines that contain inactivated or attenuated microorganisms will stimulate the innate and adaptive immune responses to a pathogen, which are both necessary for generating an adequate reserve of memory B and T cells. This goal is not achieved by protein subunits and conjugates, because they are not recognized by the Toll-like and other receptors of innate immunity (see Section 3-4). To make these proteins effective, another component, the adjuvant, a word meaning ‘helper,’ is included in the vaccine. The adjuvant activates innate immunity, producing inflammation at the vaccination site, the condition necessary to activate an adaptive immune response against the protein antigens of the vaccine.
DTaP is a widely administered vaccine that provides protection against three bacterial diseases: diphtheria, tetanus, and the whooping cough caused by Bordetella pertussis. The components of the vaccine are diphtheria toxoid, tetanus toxoid, and acellular B. pertussis. DTaP is an example of a combination vaccine that protects against more than one pathogen and disease. The B. pertussis components in the vaccine have two functions. One is to be the adjuvant that triggers the innate immune response; the other is to be the source of antigens that stimulate an adaptive immune response to B. pertussis. The inflammation induced by acellular B. pertussis ensures that effective adaptive immune responses are made against the diphtheria and tetanus toxoids.
Although research on experimental animals has identified a variety of potent adjuvants, none of them is used in human vaccines because of their toxicity. In 1924, alum—a form of aluminum hydroxide—was approved for use in human vaccines; for 73 years it was the only option. Although alum is safe, it is not a powerful adjuvant, as seen from the performance of vaccines containing diphtheria and tetanus toxoids as the antigen and alum as the adjuvant. These DT vaccines proved significantly less protective than the DTP vaccine (the precursor to the DTaP vaccine and which contained inactivated B. pertussis bacteria). Approval in 1997 of the oil-in-water adjuvant MF59 initiated a new era in adjuvant research and development, one driven by knowledge of the mechanisms by which adjuvants activate the innate immune response. Since 1997, five additional adjuvants have received approval for use in human vaccines, and others are in development (Figure 11.23). The underlying strategy is to use either microbial components recognized by Toll-like receptors or small molecules that mimic their action as the active ingredient of the adjuvant.
Figure 11.23Present and future adjuvants for use in human vaccines.
Adjuvants
Year licensed
Name
Class
Contents
In vaccines against
1924
Alum
Mineral salt
Aluminum phosphate or hydroxide
Many infectious diseases
1997
MF59
Oil-in-water emulsion
Squalene, polysorbate 80, sorbitan trioleate
Influenza
2000
Virosomes
Liposomes
Lipids, hemagglutinin
Influenza, hepatitis A
2005
AS04
Alum-absorbed TLR4 agonist
Aluminum hydroxide, monophosphoryl lipid A
Hepatitis B, human papillomavirus
2009
AS03
Oil-in-water emulsion
Squalene, polysorbate 80, α-tocopherol
Influenza
2017
AS01B
Liposome
Lipid and a natural saponin (QS-21)
Shingles (varicella zoster)
2017
CpG 1018
TLR9 agonist
Synthetic DNA composed of CpG nucleotides
Hepatitis B
In development
CpG 7909
TLR9 agonist
CpG nucleotides
Imidazoquinolines
TLR7 and TLR8 agonist
Small molecules
PolyIC
TLR3 agonist
Double-stranded RNA analogs
Pam3Cys
TLR2 agonist
Lipopeptide
Flagellin
TLR5 agonist
Bacterial protein linked to antigen
11-21Genome sequences of human pathogens have opened up new avenues for making vaccines
The original scientific strategy for vaccine development consisted of three steps: first, isolate and identify the pathogenic microorganism; second, inactivate the pathogen’s virulence while preserving its immunogenicity; and third, inject the inactivated pathogen into experimental animals as a prelude to doing the same with human subjects. These three principles of “isolate, inactivate, and inject,” first invoked by Louis Pasteur, led to the discovery during 1885–1950 of most of the vaccines that are in common use today. Subsequently, this empirical approach to vaccine development proved less fruitful. Over the past 40 years, new strategies for vaccine discovery have emerged. These approaches involve rational design and have been made possible by the technical advances of recombinant DNA, genome sequencing, and macromolecular structure determination, combined with increasing knowledge of how the immune system responds to pathogens and remembers them.
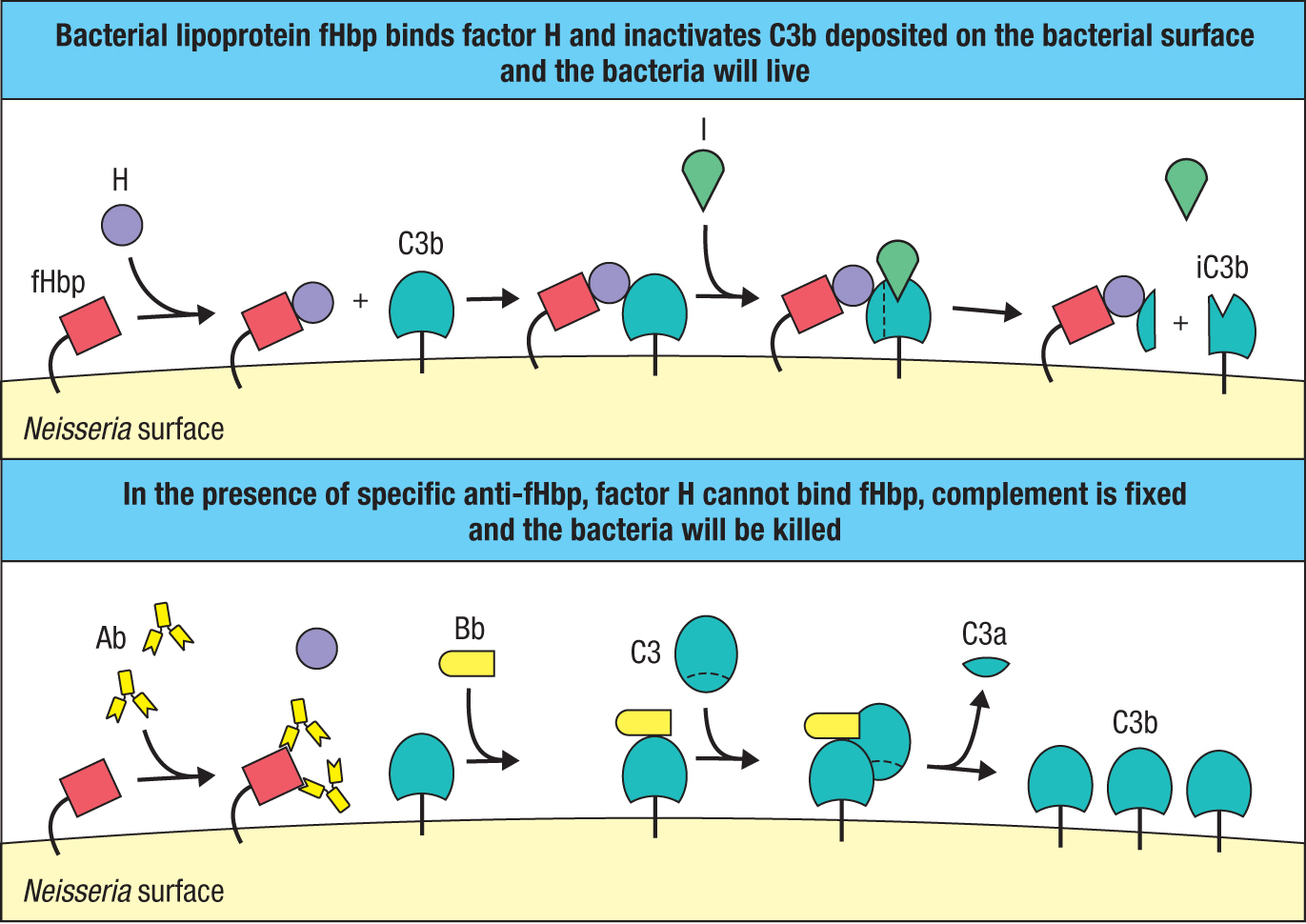
Sequencing the genomes of human pathogens has uncovered many unanticipated genes that were not detected in previous studies of their physiology and virulence. The sequences of the encoded proteins give clues to their subcellular location, function, and potential as targets for protective antibodies. An example is N. meningitidis of serogroup B. In 2000, 29 previously unknown genes were identified in this bacterium’s genome. The proteins encoded by three of the genes are highly antigenic and were combined to make Bexsero, a vaccine against meningitis B that was approved for use in Europe in 2013. All three antigenic proteins—neisserial heparin-binding protein, N. meningitidis adhesion A (Nad A), and factor H–binding protein (fHbp)—function to increase the virulence of the bacterium. The fHbp antigen is a lipoprotein expressed on the bacterial surface that binds to factor H, an inhibitor of the alternative pathway of complement activation (see Section 2-4). By coating its outer surface with this inhibitor, the pathogen fails to provoke an antibody response and cannot be killed by complement-mediated lysis (Figure 11.24). The invention and development of the Bexsero vaccine led to new knowledge of the pathogen’s physiology and its subversion of the human immune system. By contrast, the traditional approaches to vaccine development were informed and driven by preexisting knowledge of the pathogen’s physiology. To reflect this difference, the modern approach to making vaccines is called reverse vaccinology.
Figure 11.24Vaccination with Neisseria factor H–binding protein (fHbp) prevents infection from taking hold. The upper panel shows how Neisseria uses fHbp to prevent complement fixation on its surface. fHbp is a surface lipoprotein that binds factor H and uses it to inactivate any C3b deposited on the bacterial surface. Inactivation involves factor I, which cleaves C3b to give iC3b. This prevents formation of the membrane-attack complex and the lysis of Neisseria. The lower panel shows how immunization with the Bexsero vaccine, which contains fHbp, counters this evasion. Vaccination generates high-affinity IgG antibodies against fHbp that cover up its binding site for factor H on fHbp and prevent its interference with complement activation by the alternative pathway. Coating the bacteria with anti-fHbp IgG also activates the classical pathway (not shown).
11-22The rapidly evolving influenza virus requires continual vaccine development
Because influenza is an RNA virus, the replication of its genome is much more error-prone than the replication of DNA viruses such as smallpox and measles. The errors generate a diversity of mutant viruses, enabling influenza to evade human immunity; as a result, new strains of influenza spread through the human population every year. Thus immunological memory of influenza decays quickly, having a half-life of only a few years (see Section 11-6). The main targets for neutralizing antibodies are the hemagglutinin (H) and neuraminidase (N) surface glycoproteins that allow the virus to bind to cells of the respiratory epithelium and infect them. Good neutralizing antibodies block virus infection, prevent viral replication, and target virions for phagocytosis. Each winter, the new emerging strains of the virus exhibit sequence differences from the previous years’ strains in the hemagglutinin, the neuraminidase, or both. Because of these differences, several hundred million cases of influenza occur each year, resulting in around 250,000 deaths, with children and the elderly being most severely affected. To counter the evolving changes in influenza, the World Health Organization (WHO) coordinates a program to identify each year’s epidemic strains and send them to vaccine manufacturers. At some universities, for example, where large segments of the population are either young or old, the university offers a free annual ‘flu shot’ to any student, teacher, or employee who wants one. By this method, a university’s collective immunological memory of influenza is refreshed each year.
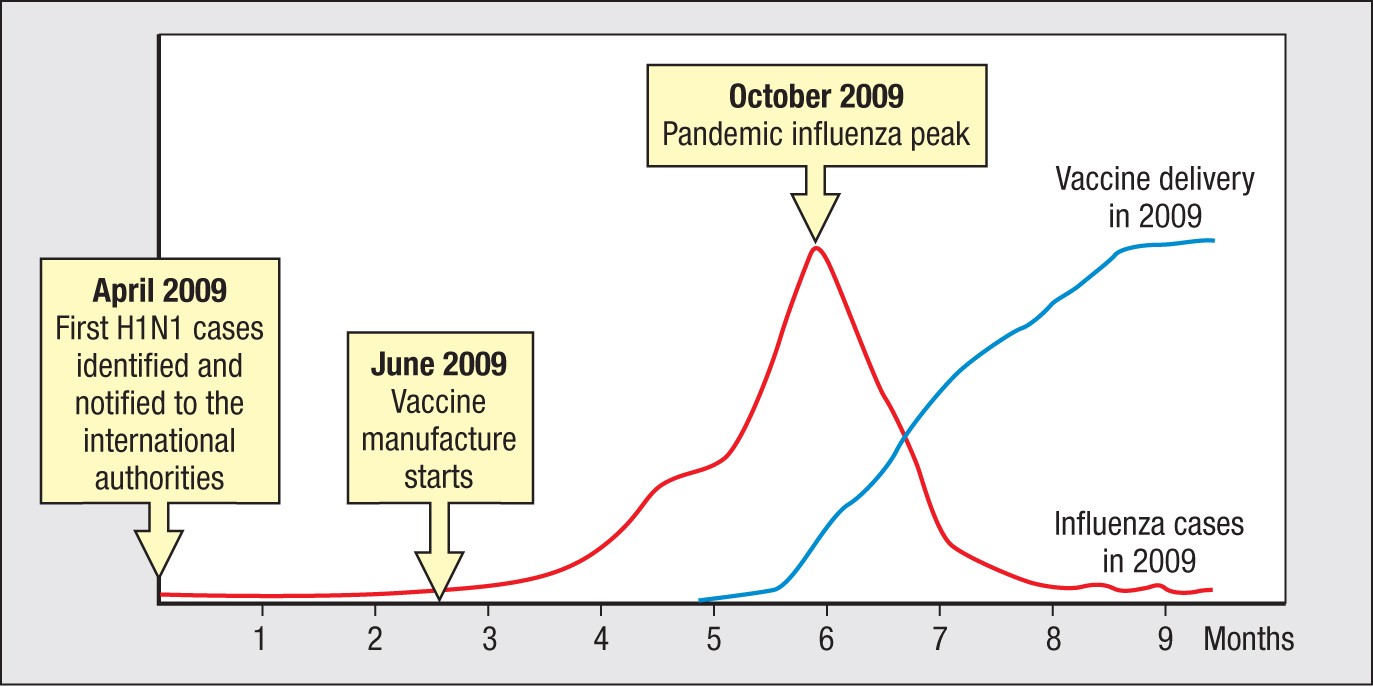
The effectiveness of the WHO program depends on timing, the goal being to vaccinate populations before the influenza arrives to cause disease and death. This requires the early detection of epidemic strains and the efficient manufacture and delivery of vaccine. In March 2009, cases of an unusually severe influenza-like illness started to appear in Mexico; by April the cause was identified as a novel H1N1 strain, and international authorities were notified. The first US case was confirmed on April 15. The hemagglutinin and neuraminidase of this virus were found to be particularly divergent, causing concern verging on alarm that a severe pandemic would occur. On June 7, manufacture of vaccine began and at the same time the influenza pandemic began to gain momentum. Unfortunately, the pandemic peaked in October 2009 (in the United States) before a vaccine was generally available (Figure 11.25). Luckily, this H1N1 did not cause severe illness or death for most of those infected, although mortality worldwide is estimated to have been in the hundreds of thousands. These events exposed the limitations of the current system for making influenza vaccines, stimulating the search for alternatives.
Figure 11.25Time course of the H1N1 influenza pandemic of 2009 and the development of a vaccine against H1N1. The red line represents the growth and decline in the number of influenza cases in the United States. The blue line represents the number of vaccine doses available against that specific strain. Data courtesy of D. Jordan/Centers for Disease Control.
Four years later, the feasibility of a bold, innovative, and provocative solution to the problem was reported. It was proposed that once a potentially epidemic strain of virus was identified and its genome sequenced, the sequence would be immediately sent electronically to manufacturers, who would synthesize the virus in one day using recombinant DNA technology. The synthetic virus would then be grown in large cultures of virus-infected cells, purified from the culture fluid, inactivated, and quality controlled. This would avoid having to grow the virus before sending it to the manufacturer. It would also avoid the traditional method of growing influenza virus in the eggs of chickens, a species that is highly susceptible to influenza. An adjuvant would be added to the vaccine so that less virus would be needed for each person vaccinated. With these changes, the protagonists proposed that a vaccine could be made within weeks, not months, after detection of a new virus, and well before any epidemic gained momentum. Although the proposal solves technological problems, the more formidable challenge is in persuading regulatory agencies to change the methods of procedure and quality control that they approve. These were put in place in the 1950s and accounted for most of the delay in making the 2009 influenza vaccine.
In the years before the COVID-19 pandemic it was generally believed that the next pandemic would be one of influenza, not of a coronavirus. Within a few days after release of the SARS-CoV-2 virus nucleotide sequence by Chinese scientists on January 13, 2020, projects to make a vaccine against SARS-CoV-2 were initiated, publicized, and rushed into development. Within 3 months some 47 vaccines using 8 different approaches were in the works (Figure 11.26). Many of these were adaptations of existing vaccines made against other viral pathogens. A first small, clinical trial of a COVID-19 vaccine was begun on March 16, 2020, and was the first of many. Because knowledge of human immune responses to SARS-CoV-2 is rudimentary, such trials are very much experiments, empirical shots in the dark, as has always been the case in search for vaccines.
Figure 11.26Candidate vaccines against SARS-CoV-2.
Type of vaccine
Number of candidate vaccines
Protein subunit
18
RNA
8
DNA
3
Nonreplicating vector
8
Replicating vector
5
Inactivated virus
2
Attenuated virus
2
Virus-like particle
1
An eventful voyage from Sydney Australia, to London, England, on SS Mooltan in 1949
In 1949 my mother was taking myself, then 6 years old, and my younger sister ‘home’ from Australia on the SS Mooltan to see our English relatives. During our voyage from Sydney to London there was an outbreak of smallpox on board the ship, although we only learned of this after dropping anchor some distance off the English coast.
An announcement was made over the public address system for all passengers to assemble in the lounge, where the captain told us “we need ‘health clearance’ and that staff from the Immigration and Quarantine Services will be coming aboard.” Small boats came alongside, and a parade of equipment-carrying officials embarked and came into the lounge. A doctor then told us that three cases of smallpox had been diagnosed and a crew member had died the previous day of the illness.
Tables were set up in the lounge in a semicircle with doctors and nurses at each. We lined up to be vaccinated. My memory is of a ‘cork’ with a needle set in its center being heated on a Bunsen burner, cooled, and then a rough circle on our upper arm was scratched and criss-crossed just piercing the skin. At the next table a small piece of gauze impregnated with a white power was placed on the scratches and rubbed in. At the third table a small dressing was applied. In time I had an incredibly painful upper arm at the vaccination site. A huge lobulated blister developed, surrounded by an extensive erythema as far as my elbow and shoulder. My whole upper arm was so grossly swollen that I had to wear a scarf as a sling. Most other passengers had the same reaction. As time went on many had burst blisters with nasty suppurating infections and high temperatures. Moving about was difficult. If someone bumped against the painful arm it could be agony and even getting dressed was difficult. Parents with babies had an especially difficult time.
Over the next few days the story of what had happened became clearer and spread throughout the ship. A Singalese crew member had become sick, but he had been hidden away for several days by his shipmates and his work shifts covered. He died of his illness, but by the time his death was reported several others were also seriously ill with high fever, delirium, and the terrible rash. A woman in a cabin along the corridor from us became ill and was taken to the sick bay. She later died. As the days passed more people succumbed. We stayed anchored far from the shore until no new cases had occurred for several days. My mother told me of a terrible fear throughout the ship and of people confined to their tiny cabins except at meal times. I am not sure how long we stayed off-shore, but before coming into port at Tilbury, a huge amount of the ship’s stock was dumped into the sea: china, glassware, cutlery, bedding, lounges, and curtains were thrown overboard. It seemed no risks were being taken. Altogether eleven people had died and were buried at sea.
On going ashore we were taken straight to my grandparents’ house. A quarantine official was assigned to visit us there every day and we were not to leave the house and garden for ten days. The man who came was a tiny, nervous fellow, who was not the least reassuring to our family. In fact he seemed quite terrified about visiting us. He would ring the front door bell and then stand three or four metres up the path away from the door to ask if we were all well. My grandparents said they found out who their friends were at this time. Some people were kind and brought food and groceries, leaving them at the front gate and asking if we needed anything else. Others would walk along the street and cross to the other side of the road until well past the house. Luckily no symptoms appeared, and after two weeks we emerged to start leading a normal life.
About two decades later as a newly graduated physiotherapist, I was a witness to the last poliomyelitis epidemic in Australia and was involved in a rehabilitation programme for the many, mostly young, people who had this dreadful disease. When I hear of opponents of vaccination, I do my best to tell them of my experiences and to urge them to do more research before refusing to vaccinate their children.
Iris Loudon
11-23The need for a vaccine and the demands placed upon it change with the prevalence of disease
In 18th-century Europe, the high probability of death or permanent facial scarring from smallpox made the risk of variolation acceptable to those who could afford it. Later, the side effects of the cowpox vaccine were tolerated during a time when smallpox still posed a threat and terrified the population. This fear persisted into living memory, as is recounted in the eyewitness account on page 329 by someone who had a brush with smallpox as a child. In the late 20th century, smallpox was eradicated and vaccination was discontinued. Smallpox vaccination was such a good preventive medicine that it put itself out of business.
Concern for a vaccine’s safety can lead to resurgence of disease, as seen for whooping cough in the 1970s. At the beginning of the 20th century, 1 in 20 of all children in the United States died from whooping cough. The DTP vaccine containing whole killed B. pertussis bacteria was introduced in the 1940s and was routinely given to infants at 3 months of age. The vaccination program produced a 100-fold decline in the annual incidence of whooping cough, from 2000 cases per million to 20 cases per million. But as people’s fear of the disease abated, their concern for the vaccine’s side effects went up. Children vaccinated with pertussis vaccine inevitably develop inflammation at the injection site, and some develop a fever that induces persistent crying. Very rarely, the vaccinated child suffers fits and either a short-lived sleepiness or a transient floppy unresponsive state, all of which cause parental anxiety. In the 1970s, awareness of these established neurological side effects was heightened by anecdotal reports of vaccination causing encephalitis and permanent brain damage, a connection that was never substantiated. Nonetheless, distrust of pertussis vaccination grew, most notably in Japan.
In Japan, DTP vaccination was introduced in 1947. By 1974, the incidence of pertussis had been reduced by more than 99%, and in that year no deaths were ascribed to the disease. In the following year, two children died soon after vaccination, raising alarm that the deaths were caused by the vaccine. During the next 5 years, the number of Japanese children being vaccinated fell from 85% to 15%, and, as a consequence, the incidence of whooping cough increased about 20-fold, as did the number of deaths from the disease. Japanese companies then developed vaccines containing antigenic proteins of pertussis instead of whole bacteria. In Japan, these acellular pertussis vaccines (DTaP) replaced the whole-cell vaccines in 1981. By 1989, the incidence of pertussis was again at the very low levels of 1974. Acellular vaccines are currently being used in other countries, because the side effects of inflammation, pain, and fever are less intense than those with DTP.
The past 20 years have seen a resurgence in the incidence of whooping cough. For example, whereas 4000 cases were reported annually in the United States during the 1980s, that number rose to 26,000 in the period from 2004 to 2010. Protection provided by acellular vaccines is thus seen to be less durable than that induced by whole-cell vaccines. Underlying this difference, DTP induces both a mucosal and a systemic immune response whereas the acellular vaccines induce only a systemic response.
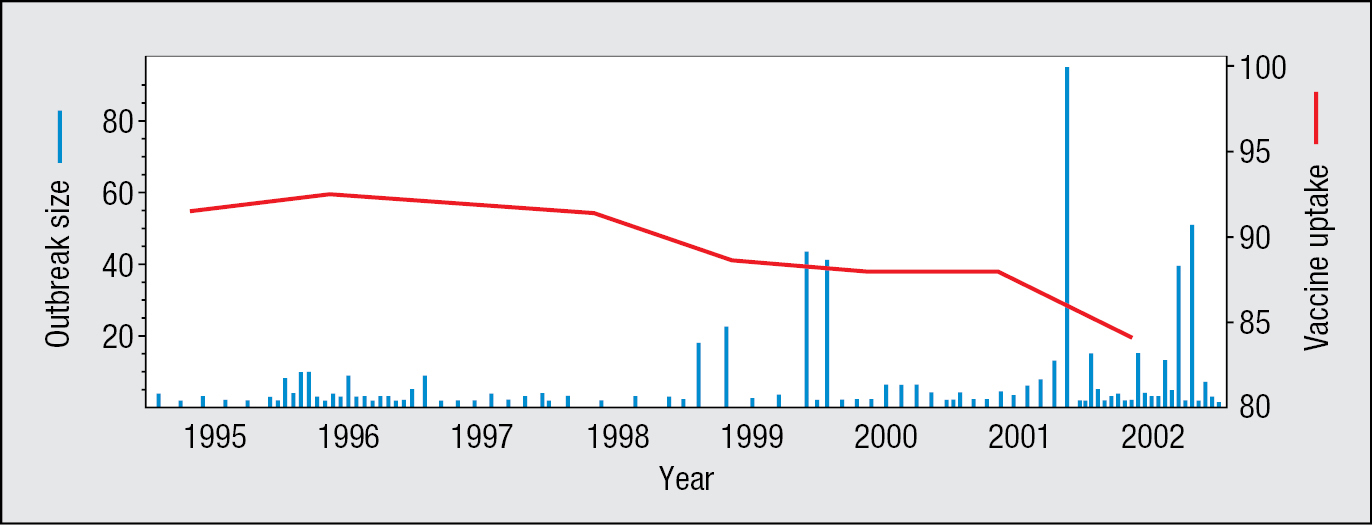
Measles is a highly infectious and potentially dangerous disease, which caused around 100 deaths per year in the United Kingdom before mass vaccination against measles began in 1968. The combined measles, mumps, and rubella (MMR) vaccine was introduced in 1988, and by the early 1990s more than 90% of children were being vaccinated against measles. Ten years after introduction of the MMR vaccine it was claimed, on the basis of 12 children diagnosed as autistic soon after vaccination, that there was a causal link between the MMR vaccine and autism. Although this claim was eventually shown to be false and its protagonist was professionally discredited, distrust of the MMR vaccine increased, causing the proportion of children being vaccinated against measles to decrease steadily, with the result that outbreaks of measles in Britain increased in size and frequency (Figure 11.27). The effects of this unfounded distrust are still being felt. In spring 2013, an outbreak of more than 1000 cases of measles occurred in Swansea, a city where more than 6000 children had never received the MMR vaccine.
Figure 11.27As fewer children were vaccinated against measles virus, the incidence of measles infections increased. Data are from the UK population. Since 1998, when a link between the MMR vaccine and autism was falsely asserted, the number of children vaccinated (red line) decreased and outbreaks of measles (blue bars) increased in both frequency and size. The vaccine uptake is the percentage of children completing a primary course of the MMR vaccine at their second birthday. Data courtesy of V.A.A. Jansen and M.E. Ramsey.
In 1998, the vast majority of the UK population had protective immunity against measles virus as a result of previous vaccination or infection. In this situation, a population has a herd immunity, which indirectly protects the minority of unvaccinated people. The pathogen fails to create an epidemic, because of the low probability of finding susceptible individuals and creating a chain of infection. If the proportion of children being immunized against measles continues to decrease, however, the proportion of the UK population with no immunity to measles could become sufficiently large for herd immunity to be lost. An outbreak of measles could then more easily become an epidemic. Although measles was declared eliminated from the US population in 2000, more than 1280 cases of measles were reported there in 2019, and a further 12 up to May 2020.
11-24Vaccines have yet to be made against pathogens that establish chronic infections
The diseases for which current vaccines are effective are acute infections, which resolve in a matter of weeks, either by elimination of the pathogen or the death of the human host (Figure 11.28). As pathogens cannot survive if they eliminate too many of their hosts, a majority of people do survive infection with the common, familiar pathogens. At its worst, smallpox killed only one-third of the people it infected. Vaccines that protect against these diseases work because they stimulate immune responses that mimic the protective response provoked by natural infection with the pathogen.
Figure 11.28Diseases for which vaccines are available. Not all of these vaccines are equally effective, and not all are in routine use.
Available vaccines for infectious diseases of humans
Bacterial diseases
Types of vaccine
Viral diseases
Types of vaccine
Diphtheria (Corynebacterium diphtheriae)
Toxoid
Yellow fever
Attenuated virus
Tetanus (Clostridium tetani)
Toxoid
Measles
Attenuated virus
Pertussis (Bordetella pertussis)
Killed bacteria.
Subunit vaccine composed of pertussis toxoid and other bacterial antigens
Mumps
Attenuated virus
Paratyphoid fever (Salmonella paratyphi)
Killed bacteria
Rubella
Attenuated virus
Typhus fever (Rickettsia prowazekii)
Killed bacteria
Polio
Attenuated virus (Sabin) or killed virus (Salk)
Cholera (Vibrio cholerae)
Killed bacteria or cell extract
Varicella (chickenpox) Varicella (shingles)
Attenuated virus
Plague (Yersinia pestis)
Killed bacteria or cell extract
Influenza
Inactivated virus
Tuberculosis (Mycobacterium tuberculosis)
Attenuated strain of bovine Mycobacterium tuberculosis (BCG)
Rabies
Inactivated virus (human).
Attenuated virus (dogs and other animals).
Recombinant live vaccinia-rabies (animals)
Typhoid fever (Salmonella typhi)
Vi polysaccharide subunit vaccines.
Live-attenuated oral vaccine
Hepatitis A
Subunit vaccine (recombinant hepatitis antigen)
Meningitis (Neisseria meningitidis)
Purified capsular polysaccharide
Hepatitis B
Subunit vaccine (recombinant hepatitis antigen)
Bacterial pneumonia (Streptococcus pneumoniae)
Purified capsular polysaccharide.
Polysaccharide conjugated to protein
Human papillomavirus
Subunit vaccine (virus coat proteins)
Meningitis (Haemophilus influenzae)
H. influenzae polysaccharide conjugated to protein
Rotavirus
Attenuated virus.
Recombinant live virus
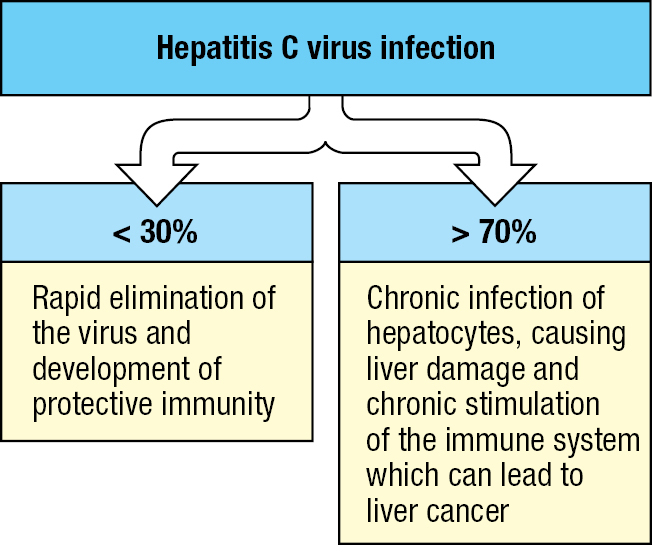
Figure 11.30A minority of people exposed to hepatitis C virus defeat the virus, while the majority develop chronic infection.
By contrast, devising vaccines that work against chronic infectious diseases has proved difficult (Figure 11.29). After infection, pathogens such as the malarial parasite Plasmodium falciparum, the bacterium Mycobacterium tuberculosis, and the human immunodeficiency virus (HIV) modify human immune systems to make them work in their favor. From studying patients with these conditions, it is not always clear what type of immune response would be effective if triggered by a vaccine. In this context, hepatitis C virus (HCV) is informative because it causes both acute and chronic infections. Depending on the population, 15–30% of infected people suffer mild disease symptoms and make a primary immune response that clears the infection, leaving an immunological memory of the virus (Figure 11.30). Nor does the acute infection ever kill the human host. By contrast, the majority of people infected with HCV, some 70 million worldwide, make an ineffective immune response that permits chronic infection and persisting disease in which the infected liver goes through cycles of cellular destruction and tissue regeneration. This disease progression can lead to cancer or liver failure and the need for a liver transplant, for which people chronically infected with HCV comprise most of the applicants. But despite many attempts to make an HCV vaccine, that goal has yet to be achieved.
Figure 11.29Diseases for which better vaccines are needed.‡ The measles vaccines currently used are effective, but they are heat sensitive and require carefully controlled refrigeration, as well as reconstitution, before use. In some tropical countries this reduces their usefulness. The BCG vaccine against tuberculosis has limited effect and is not included in routine vaccination schedules. § Estimated mortality from the effects of chronic infection. Sources of estimated mortality, incidence, and prevalence data: World Malaria Report 2019 (WHO: 2019); UNAIDS 2018 Global HIV Statistics (UNAIDS: 2019); other figures from World Health Organization estimates for 2017–2018.
Some diseases for which effective vaccines are not yet available
Disease
Estimated global annual mortality
Estimated global annual incidence* or total number of people infected†
Malaria
405,000
228 million*
Schistosomiasis
23,000
200 million†
Tuberculosis‡
1.3 million
10 million*
Diarrheal disease
1.7 million
1.7 billion*
HIV/AIDS
770,000
1.7 million*
37.8 million†
Measles‡
110,000
20 million*
Hepatitis C
339,000§
34 million*
71 million†
However, in the past decade there has been considerable success in the treatment of people with chronic HCV using drugs that inhibit viral replication. An 8- to 12-week treatment of taking 1–3 pills a day can eliminate the virus, and this is considered a ‘virologic’ cure if the virus remains undetectable 3 months after the treatment. Some 5 million people with chronic HCV have been treated with these drugs.
11-25Vaccine development faces greater public scrutiny than does drug development
The development of a vaccine is inherently more challenging than the development of a drug. One difference is that drugs are given to sick people who tend to be grateful for any alleviation of their condition, whereas vaccines are administered to healthy individuals, often the very young, for whom immediate adverse affects of vaccination can be of greater concern to their parents than potential long-term benefits. Another difference is that drugs are prescribed on a case-by-case basis, whereas vaccination is usually part of a large program mandated by an authority and applied to the population at large. Exemplifying the latter is the worldwide vaccination program aimed at eradicating polio. Most people experience and appreciate the beneficial effects of therapeutic drugs. By contrast, most people who have been vaccinated against smallpox, polio, and the other infections that historically killed many people have never seen or experienced the disease, so cannot appreciate what horrors they have missed. The only situation in which a vaccine’s benefit can truly be felt is when exposure is obvious, because unvaccinated members of the community are suffering and succumbing to disease.
Drugs and vaccines all have unwanted side effects, for which risk must be weighed against benefit. Because a vaccine’s side effects might cause symptoms in healthy children, the standards set for vaccines are always higher than those for drugs. This sensitivity has also led to periodic episodes when disease or other syndromes are erroneously and irresponsibly attributed to be the consequence of vaccination, as autism and the MMR vaccine were linked in 1998 (see Section 11-23). Because of the nature of herd immunity, individuals can gain the benefit of mass vaccination programs without sharing the risk. This is yet another difference from drugs, for which the cost and the benefit are inseparable. However, if enough individuals take this selfish strategy, then herd immunity will be lost, and in the ensuing epidemic they too could lose out.
Although vaccines are the most frequent and successful of medical interventions, their acceptance and use has been greatly affected by factors having little to do with science or medicine. For example, an early version of a rotavirus vaccine was abandoned in 1999 because of its side effect, a bowel obstruction that, if not treated with an enema or surgery, could be fatal, which reportedly occurred in 1 of 7000 children vaccinated. Although the decision to abandon the vaccine could be justified in the United States, where only 1 in 100,000 deaths is caused by rotavirus infection, the vaccine would have greatly benefited the children of other countries, such as India (see Figure 11.20), where as many as 1 in 200 of all children were dying from rotavirus infection. Consequently, the decision to discontinue rotavirus vaccination in some countries led to many more deaths from infection than lives saved by abandoning vaccination. Fortunately, there are now safe and effective rotavirus vaccines (see Section 11-17).
Summary
Vaccination is preventive medicine. It involves the deliberate immunization of healthy people with some form of a pathogen or its component antigens and induces a protective immunity that prevents any future infection from causing disease. Vaccines consist of killed whole pathogens, live-attenuated strains, nonpathogenic species related to the pathogen, or macromolecules of the pathogen that are secreted or present on the pathogen’s surface. Vaccination has saved millions of lives and reduced the incidence of many common infectious diseases, particularly in the rich industrialized countries. The prevention of infectious disease by vaccination demonstrates how manipulation of the immune response can improve the health of human individuals and their communities. The pathogens for which effective vaccines have already been found are those that cause acute infections and are not highly mutable. When these pathogens cause epidemic disease, many people survive and with increased immunity, showing how human immune systems can respond to the infection in helpful ways. Death, when it occurs, is because the immune response was too slow. What vaccination achieves is to prime the immune response ahead of infection, giving the human host an edge on the pathogen. By reducing the incidence of disease, successful vaccination programs inevitably lead to decreased public awareness of the effects of disease, which combines with increasing worry about the safety and possible side effects of the vaccination.
Until the approval of the hepatitis B virus subunit vaccine in 1986, vaccine development was largely a process of trial and error, one in which a knowledge of immunological mechanisms played little part and the guiding principle was for the vaccine to resemble the natural pathogen as closely as possible. Although this approach worked well for pathogens causing acute infections, it failed to discover vaccines against pathogens that establish chronic infections and cause chronic disease. In people with these diseases, the immune response has not been helpful, and the challenge is to find out what would be helpful. Because of the risks and costs inherent to vaccine development, as well as the political and legal complications associated with their side effects, both real and imagined, commercial vaccine manufacture was a declining industry for more than 40 years. From 1967 to 2019 the number of vaccine manufacturers in the United States declined from 26 to 7. The future is looking brighter, because of the application of recombinant DNA technology. This revolution started in the 1980s, and was then expanded and refined to provide the high-throughput analyses required by the Human Genome Project and its successors. This in turn has led to reverse vaccinology, by which a pathogen’s genome is mined to identify candidate antigens that are then studied to determine their biology and how they interact with the immune system. This approach increases our basic understanding of host–pathogen relationships, while at the same time engages in its application: the discovery of new vaccines. Increasing knowledge of innate immunity is leading to new adjuvants—the natural ligands of Toll-like receptors and other sensors of innate immunity. On the horizon is the possibility of accelerated approaches for making vaccines against emerging strains of influenza, so that the world’s populations can be vaccinated before the virus arrives.
historical procedure for immunization against smallpox in which a small amount of live smallpox virus was introduced through scarification of the skin. (Chapter 11)
a close relative of the cowpox virus. It causes a limited infection in humans that leads to immunity to the human smallpox virus. Was used as the vaccine against smallpox that led to eradication of the disease. (Chapter 11)
a vaccine containing viral particles that have been deliberately killed by heat, chemicals, or radiation. Also called inactivated virus vaccine. (Chapter 11)
a vaccine composed of live virus particles that have an accumulation of mutations that impede their growth in human cells and capacity to cause disease. (Chapter 11)
cytotoxic protein secreted by the bacterium Corynebacterium diphtheriae, the cause of diphtheria, and which causes the disease symptoms. The diphtheria vaccine consists of an inactive form of the toxin called diphtheria toxoid. (Chapter 11)
vaccine made from capsular polysaccharides bound to an immunogenic protein such as tetanus toxoid. The protein provides peptide epitopes that stimulate CD4 T cells to help B cells specific for epitopes of the polysaccharide. (Chapter 11)
an innovative approach to vaccine development that discovers and uses new knowledge of the physiology of the pathogen and how it exploits the human immune system. (Chapter 11)
the phenomenon whereby those people in a population who have no protective immunity against a pathogen are largely protected from infection. This occurs when the majority of the population has protective immunity (usually as a result of vaccination) and is resistant to the pathogen. (Chapter 11)