2.4 Fixation and Staining for Bright-Field Microscopy
For bright-field microscopy, we can increase detection and resolution by fixation and staining. Fixation and staining are procedures that usually kill the cell. Fixation is a process by which cells are made to adhere to a slide in a fixed position. We can fix cells with methanol or by heat treatment to denature the cell’s proteins, whose exposed side chains then adhere to the glass. Staining is the use of a molecule that absorbs much of the incident light, usually over a wavelength range that results in a distinctive color (Fig. 2.21). The use of chemical stains was developed in the nineteenth century, when German chemists used organic synthesis to invent new coloring agents for clothing. Clothing was made of natural fibers such as cotton or wool, so a substance that dyed clothing would be likely to react with biological specimens.
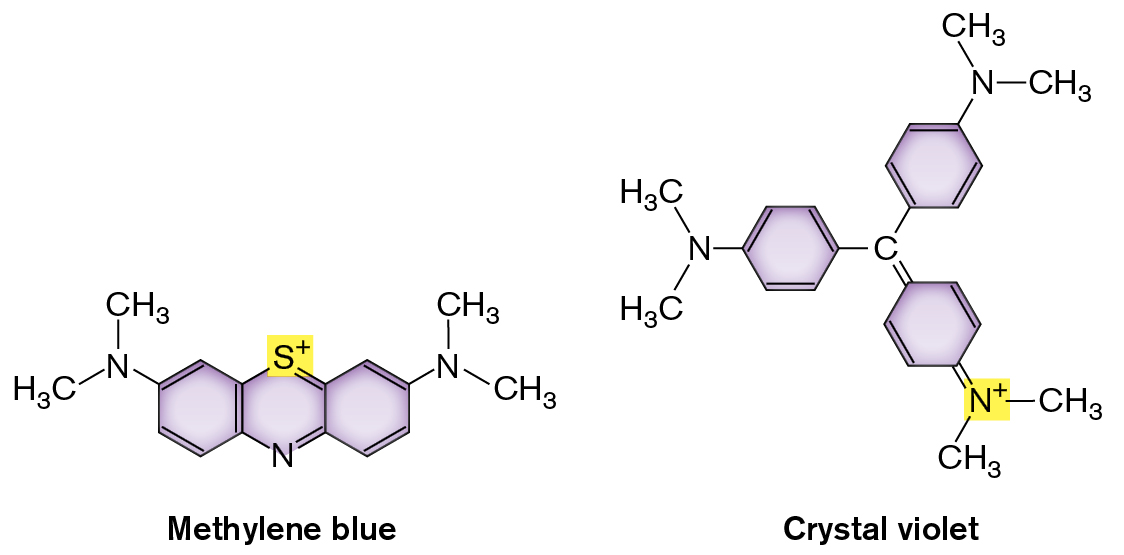
How do stains work? Most stain molecules contain conjugated double bonds or aromatic rings that absorb visible light (Fig. 2.21). Stains also have positively charged groups that bind cell-surface components with negative charge, such as the phosphoryl groups of membrane phospholipids (discussed in Section 3.2). Different stains vary with respect to the strength of their binding and the degree of binding to different parts of a cell.
More information
Two molecular structures of Methylene blue and Crystal violet are shown. The structure of Methylene blue consists of three benzenes bonded. C 1 at the top of the center benzene is substituted by a sulfur plus ion, and C 4 at the bottom is substituted by nitrogen. C 6 at the top left of the left benzene is single bonded to nitrogen, and nitrogen is single bonded to two methyl groups. C 2 of the right benzene is single bonded to nitrogen, and nitrogen is single bonded to two methyl groups. The structure of Crystal violet consists of a central carbon bound to three benzene rings. In the benzene ring at the left, C 6 is single bonded to a nitrogen which is single bonded to two methyl groups. The C 3 is single bonded to the central carbon. In the benzene ring at the top, C 6 of the benzene is single bonded to nitrogen, and the nitrogen is single-bonded two methyl groups. The C 3 is single bonded to the central carbon. In the benzene ring at the bottom, C 4 is double-bonded to a nitrogen plus ion which is single bonded to two methyl groups. The C 1 is double bonded to the central carbon.
FIGURE 2.21 ■Chemical structure of stains. Methylene blue and crystal violet are cationic (positively charged) dyes.
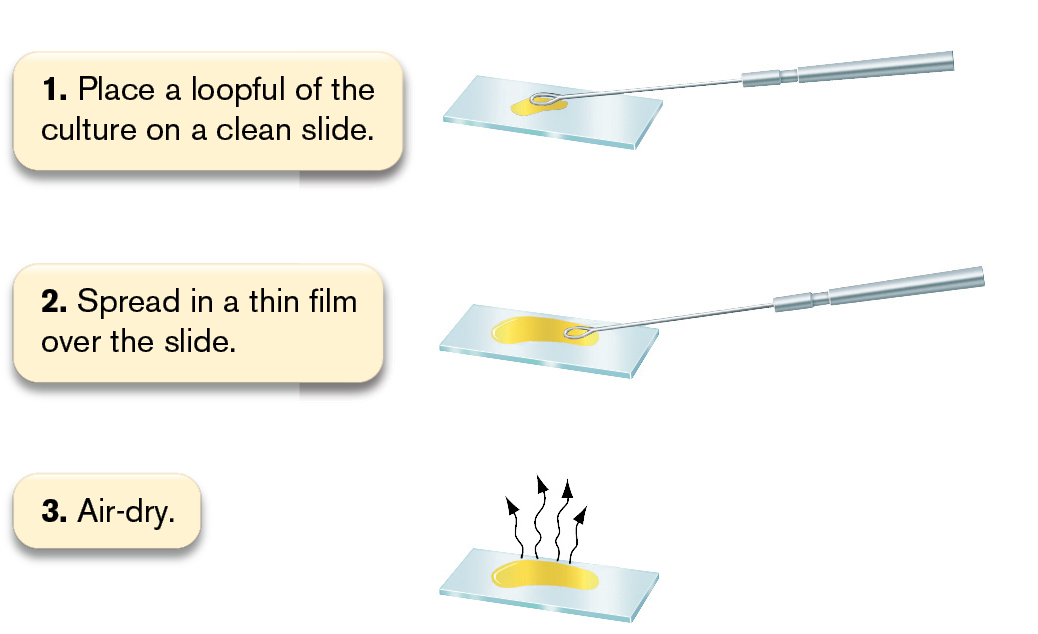
Simple stains. A simple stain adds dark color specifically to cells, but not to the external medium or surrounding tissue (in the case of pathological samples). The most commonly used simple stain is methylene blue (see Fig. 2.6E), originally used by Robert Koch in the nineteenth century to stain bacteria. A typical procedure for fixation and staining is shown in Figure 2.22. First we fix a drop of culture on a slide by treating it with methanol or by heating it on a slide warmer (steps 1–4 in the figure). Either of these treatments denatures cell proteins, exposing side chains that bind to the glass. We then flood the slide with methylene blue solution (step 5). The positively charged molecule binds to the negatively charged cell envelope of fixed bacteria. After excess stain is washed off and the slide has been dried, we observe it under high-power magnification using immersion oil (steps 6–8).
More information
An illustration shows 8 steps involved in the process of methylene blue staining.
Step 1: Place a loopful of the culture on a clean slide. The illustration shows bacterial culture placed on a slide using a metal loop.
Step 2: Spread in a thin film over the slide. The illustration shows the culture spread thin using the loop.
Step 3: Air-dry. The illustration shows the culture drying.
More information
An illustration shows 8 steps involved in the process of methylene blue staining.
Step 4: Fix cells to slide by adding a drop of methanol, then air-dry. The illustration shows methanol added to the slide using a dropper.
Step 5: Stain with methylene blue for 1 minute. The illustration shows blue stain added to the slide using a dropper.
More information
An illustration shows 8 steps involved in the process of methylene blue staining.
Step 6: Wash off the stain with water. The illustration shows the slide held at an angle, with water added to remove excess stain.
More information
An illustration shows 8 steps involved in the process of methylene blue staining.
Step 7: Blot off excess water. The illustration shows the stained slide between two sheets of blotting paper.
More information
An illustration shows 8 steps involved in the process of methylene blue staining.
Step 8: View under the microscope. The illustration shows the stained slide on a microscope stage.
FIGURE 2.22 ■Procedure for simple staining with methylene blue.
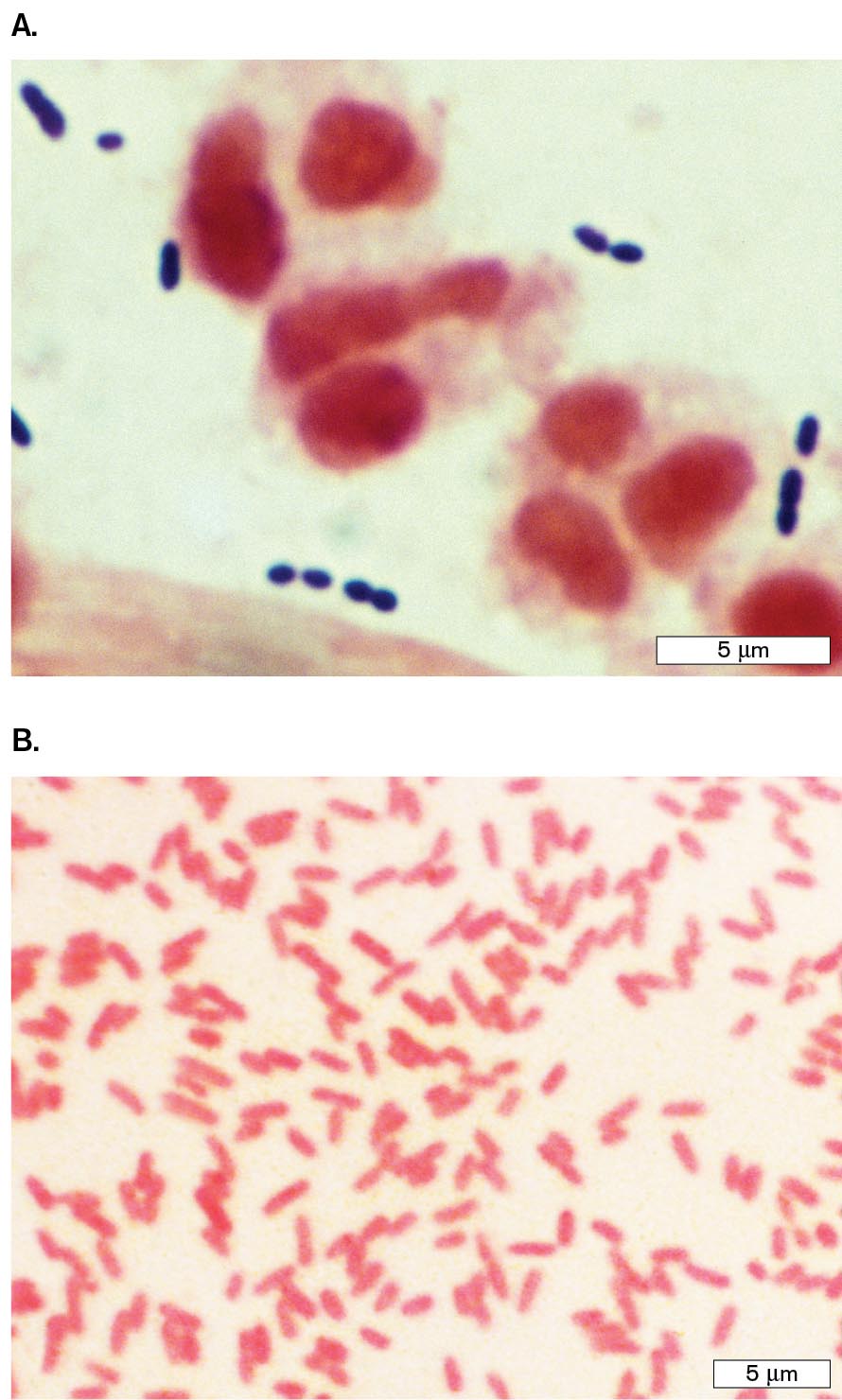
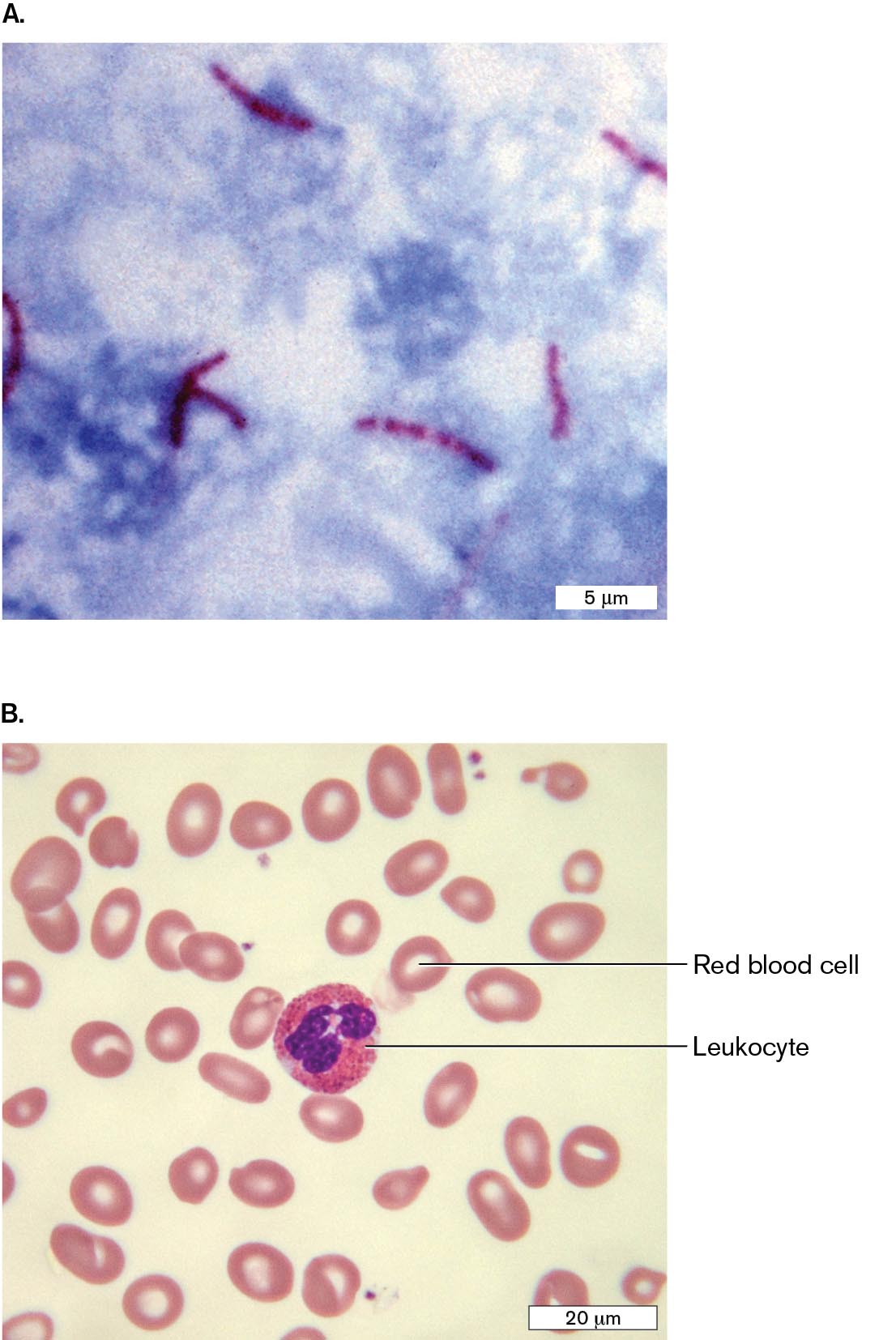
Differential stains. A differential stain colors one kind of cell but not another. The most famous differential stain is the Gram stain, devised in 1884 by the Danish physician Hans Christian Gram (1853–1938). Gram first used this stain to distinguish pneumococcal bacteria (Streptococcus pneumoniae) from human lung tissue. In Figure 2.23A, Gram-stained S. pneumoniae bacteria appear dark purple among unstained white blood cells. Other species of bacteria, such as Proteus mirabilis (a cause of urinary infections), fail to retain the purple stain (Fig. 2.23B). Different bacterial species are classified as Gram-positive or Gram-negative, depending on whether they retain the stain. Members of the phylum Proteobacteria (Pseudomonadota) stain Gram-negative, whereas most of the Firmicutes (Bacillota) stain Gram-positive. Other taxa may stain Gram-negative or Gram-variable.
More information
Two micrographs show examples of Gram-positive and Gram-negative bacteria.
A micrograph of diplococci Gram-positive Streptococcus pneumoniae bacteria. The bacteria are each about 0.5 micrometer in diameter and are stained purple. There are much larger red ovals in the specimen too.
A micrograph of rod shaped Gram-negative Proteus mirabilis bacteria. The bacteria are each about 1 micrometer in length and 0.25 micrometer in width. The bacteria are stained pink.
FIGURE 2.23 ■Gram staining of bacteria.A. Gram stain of a sputum specimen from a patient with pneumonia, containing Gram-positive Streptococcus pneumoniae (purple diplococci) among white blood cells in pus. The white blood cell nuclei stain pink (counterstain). B. Gram-negative Proteus mirabilis (pink rods).CDC/DR. MIKE MILLERCDC/SCIENCE SOURCE
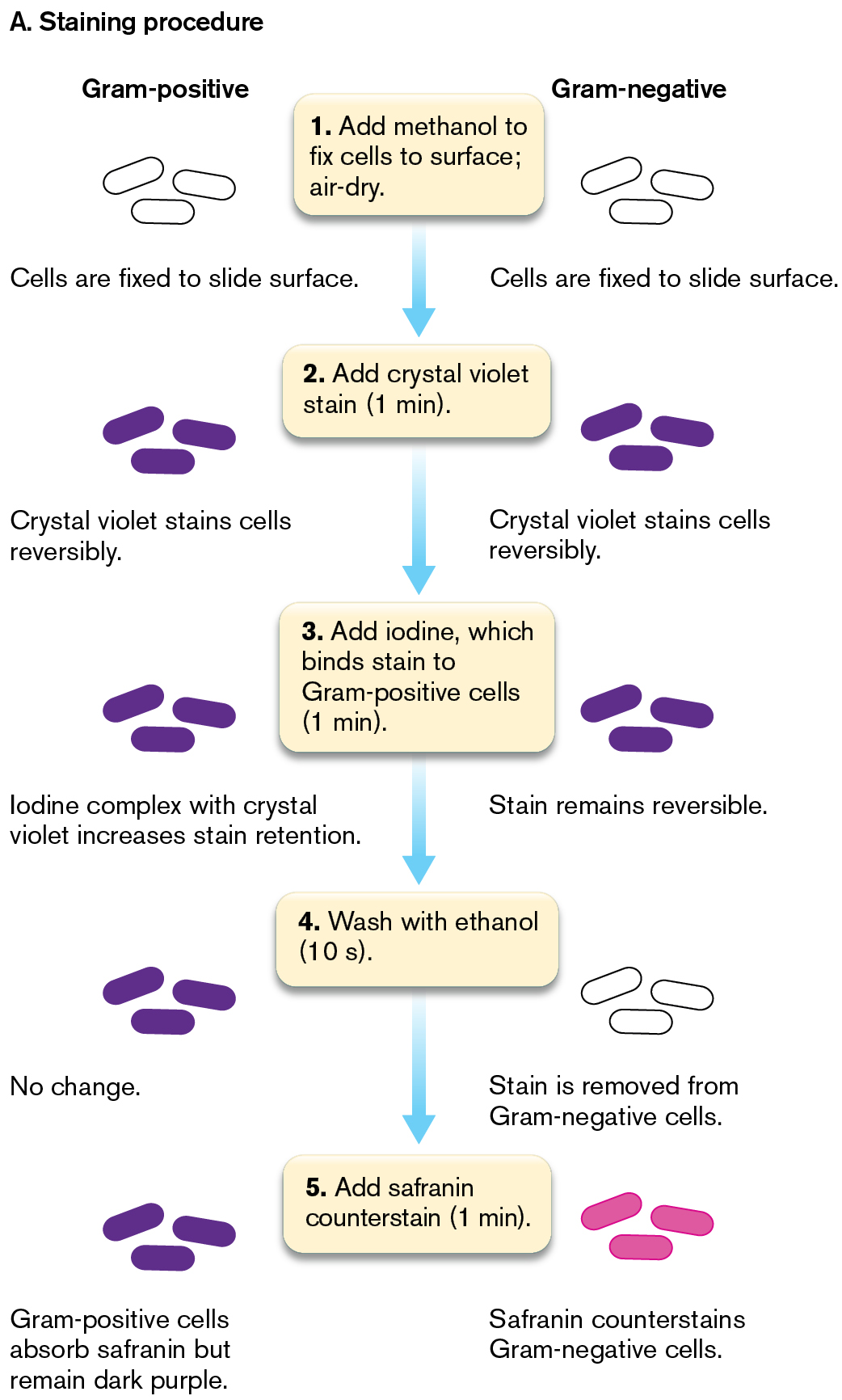
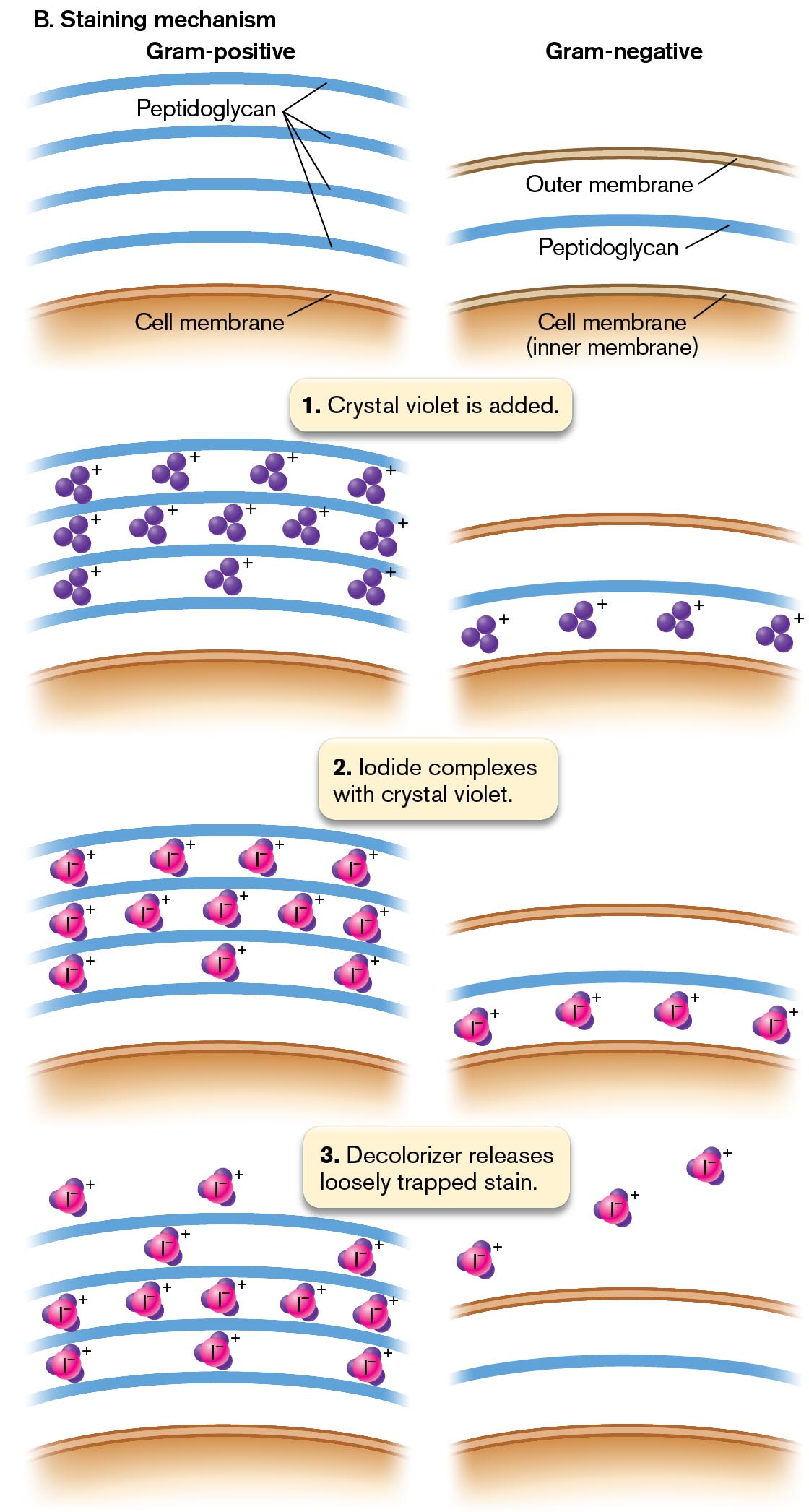
In the Gram stain procedure (Fig. 2.24A), a dye such as crystal violet binds to the bacteria; it also binds to the surface of human cells, but less strongly. After the excess stain is washed off, we apply a mordant, or binding agent. The mordant used is iodine solution, which contains iodide ions (I−). The iodide complexes with the positively charged crystal violet molecules trapped inside the cells (step 3 in the figure). The crystal violet–iodide complex is now held more strongly within the cell wall. The thicker the cell wall, the more crystal violet–iodide molecules are held.
Next we add a decolorizer, ethanol, for a precise time interval (typically 10 seconds). The decolorizer removes loosely bound crystal violet–iodide, but Gram-positive cells retain the stain tightly (Fig. 2.24A, step 4). The Gram-positive cells that retain the stain appear dark purple, while the Gram-negative cells are colorless. Timing the decolorizer step is critical because if it lasts too long, the Gram-positive cells, too, will release their crystal violet stain. In the final step, a counterstain, safranin, is applied (step 5). This process allows the visualization of Gram-negative material, which is stained pale pink by the safranin. Gram-positive cells also retain safranin; thus, if the cells are decolorized too long, both Gram-positive and Gram-negative cells will appear pink because of the safranin.
How does the Gram stain distinguish different cell types? Most Gram-negative species of bacteria possess a cell wall that is thinner and more porous than that of Gram-positive species (discussed in Chapter 3). A Gram-negative cell wall has only one to three layers of peptidoglycan (sugar chains cross-linked by peptides), whereas a Gram-positive cell has five or more layers (Fig. 2.24B). The multiple layers of peptidoglycan retain enough stain complex that the cell appears purple.
More information
Two illustrations show the principles of Gram staining for Gram-positive and Gram-negative bacteria.
The first illustration shows the steps of the Gram-staining procedure. The appearance of Gram-positive and Gram-negative bacteria are shown at each step. Step 1: Add methanol to fix cells to the surface of the slide and air-dry. At this step, both Gram-positive and Gram-negative bacteria appear colorless.
Step 2: Add crystal violet stain for 1 minute. At this step, both Gram-positive and Gram-negative bacteria appear purple. The cells are stained with crystal violet reversibly.
Step 3: Add iodine for 1 minute, which binds the stain to gram-positive cells. At this step, both Gram-positive and Gram-negative bacteria continue to appear purple. In Gram-positive cells, iodine forms a complex with crystal violet which increases retention of the crystal violet stain. In Gram-negative cells, the crystal violet stain remains reversible.
Step 4: Wash with ethanol for 10 seconds. At this step, the Gram-positive bacteria appear purple, there is no change to stain retention in these cells. The Gram-negative bacteria appear colorless, as the stain is removed from these cells. Step 5: Add safranin counterstain for 1 minute. At this step, the Gram-positive bacteria appear purple and the Gram-negative bacteria appear pink. The Gram-positive cells do absorb safranin, but they remain a dark purple color. The Gram-negative cells are counterstained pink by the safranin.
More information
The second illustration shows the mechanism of Gram-staining. Two columns show the cell membrane and peptidoglycan layers in Gram-positive and Gram-negative bacteria. In Gram-positive cells, there are several peptidoglycan layers beyond the cell membrane. In Gram-negative cells a single peptidoglycan layer is present between the outer membrane and the inner cell membrane. During the first step of Gram-staining, crystal violet is added. In Gram-positive cells, positively charged crystal violet ions are present between the layers of peptidoglycan. In Gram-negative cells, positively charged crystal violet ions are present between the peptidoglycan layer and the inner cell membrane. During the second step of Gram-staining, iodine complexes with crystal violet. In Gram-positive cells, positively charged iodine complexes are present between the peptidoglycan layers. In Gram-negative cells, positively charged iodine complexes are present between the peptidoglycan layer and the inner cell membrane. During the third step, decolorizer releases loosely trapped stain. In Gram-positive cells, the majority of iodine complexes remain between the peptidoglycan layers. In Gram-negative cells, the iodine complexes are shown beyond the outer cell membrane.
FIGURE 2.24 ■The Gram stain.A. Gram-positive cells have thick cell walls that retain the crystal violet stain. Gram-negative cells have thinner cell walls, which lose the crystal violet stain but are counterstained by safranin. B. In a Gram-positive cell, multiple layers of cell-wall peptidoglycan retain the crystal violet–iodide complex. In a Gram-negative cell, the stain leaks out.
The Gram stain became a key tool for identifying pathogens in the clinical laboratory. As we’ll see in Chapter 3, the Gram stain effectively distinguishes Proteobacteria (a diverse group of Gram-negative bacteria with a thin cell wall and an outer membrane) from Firmicutes (Gram-positive bacteria with a thick cell wall and no outer membrane). Proteobacteria include Escherichia coli and many related intestinal bacteria. Another phylum that stains Gram-negative is Bacteroidetes (Bacteroidota), which work with Proteobacteria to digest our food (discussed in Chapters 13 and 21).
Our colon also contains Gram-positive Firmicutes such as species of Clostridium and Enterococcus. Most intestinal bacteria are mutualists; that is, they share positive contributions with their host (the human body). However, the gut community may be invaded by deadly pathogens, such as the pathogenic Escherichia coli strain O157:H7 or the Gram-positive Enterococcus faecalis and Clostridioides difficile.
Still other groups of bacteria and archaea have different kinds of cell walls that may stain Gram-positive, Gram-negative, or variable (discussed in Chapters 18 and 19). Moreover, even Firmicutes such as Bacillus species show variable stain results depending on their growth state and environmental conditions.
Other differential stains reveal components specific to certain classes of bacteria (Fig. 2.25):
More information
Two micrographs show examples of acid-fast stain and Giemsa stain as seen with light microscopy.
A micrograph of Mycobacterium tuberculosis stained with acid-fast stain. The M. tuberculosis cells are rod-shaped and stained red. The cells are each about 5 micrometers long. The rest of the slide has a blue tint.
A micrograph of human blood cells stained with Giemsa stain. Light red biconcave cells are identified as red blood cells. A leukocyte, or white blood cell, is identified. The leukocyte contains granules that are stained bright red and a segmented nucleus stained dark purple.
FIGURE 2.25 ■Differential stains.A. Acid-fast stain of Mycobacterium tuberculosis (red) in sputum. B. Giemsa stain of human blood film.CDC/DR. GEORGE P. KUBICANATURE’S FACES/SCIENCE SOURCE
Acid-fast stain (Ziehl-Neelsen). Carbolfuchsin specifically stains mycolic acids of Mycobacterium tuberculosis and M. leprae, the causative agents of tuberculosis and leprosy, respectively (Fig. 2.25A).
Giemsa stain for blood film. A mixture of methylene blue, eosin (pink, protein-binding), and Azure B (blue) is used to stain blood cells and associated parasites. Figure 2.25B shows red blood cells appearing pink, and a monocyte and a leukocyte whose nuclei stain purple.
Negative stain. A negative stain is a suspension of opaque particles such as India ink added to darken the surrounding medium and reveal transparent components such as the outer capsule of a pathogen (presented in Chapter 3). Other kinds of negative stains are used for electron microscopy (see Section 2.6).
Antibody stains. Stains linked to antibodies can identify precise strains of bacteria or even specific molecular components of cells. The antibody binds a specific cell protein. The antibody may be “conjugated” (attached) to a reactive enzyme for detection or to a fluorophore (fluorescent molecule) for immunofluorescence microscopy. Fluorescence microscopy is discussed next, in Section 2.5.
To Summarize
Fixing and staining a specimen kills it but improves contrast and resolution.
A differential stain colors one kind of cell but not another.
The Gram stain differentiates between two major bacterial taxa, which stain either Gram-positive (Firmicutes) or Gram-negative (Proteobacteria). Human tissues stain Gram-negative. Other bacteria and archaea may stain Gram-negative or Gram-variable.
Giemsa stain distinguishes different kinds of blood cells and parasites.
Antibody stains detect a very specific cell type or component. The antibody is conjugated to an enzyme or a fluorophore.
A differential stain that distinguishes cells that possess a thick cell wall and retain a positively charged stain (Gram-positive) from cells that have a thin cell wall and outer membrane and fail to retain the stain (Gram-negative).
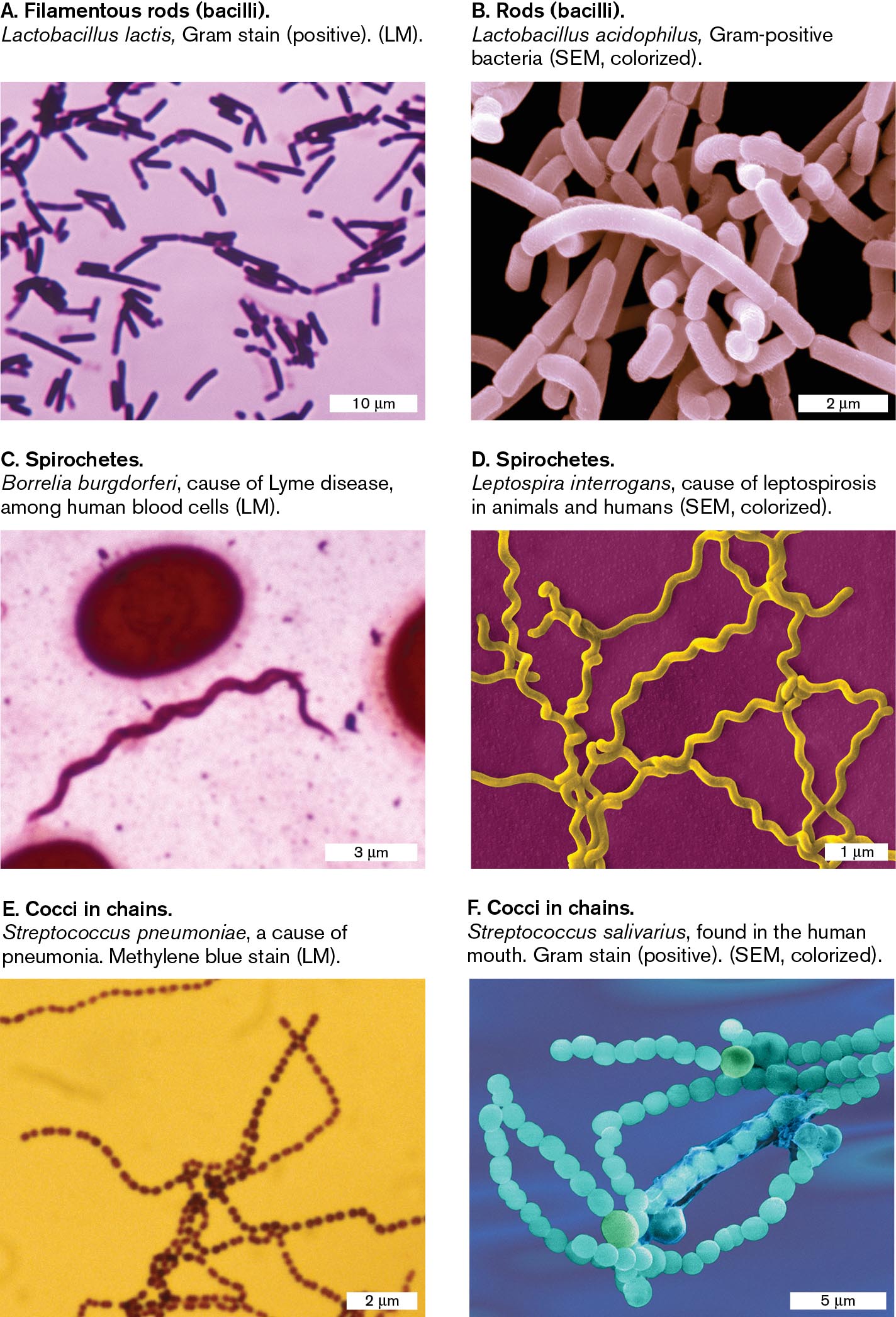
Six micrographs show examples of the common shapes seen in bacterial cells. The first micrograph shows filamentous rods. The second micrograph shows rods. The third and fourth micrographs shows spirochetes. The fifth and sixth micrographs show cocci in chains.
A micrograph of Lactobacillus lactis, a filamentous rod shaped bacteria, as seen under a light microscope. There are many rod-shaped bacteria, each 3-5 micrometers long and 1 micrometer wide. The cells are stained purple.
A micrograph of Lactobacillus acidophilus, a rod shaped bacterium, as seen with a scanning electron microscope. A group of rod shaped bacteria are clumped together. The bacterial cells are each about 2 micrometers long and 0.5 micrometer wide.
A micrograph of Borrelia burgdorferi, a spirochete shaped bacterium, as seen under a light microscope. A single spiral shaped bacterium is on the slide. The cell is about 15 micrometers long and 0.25 micrometer wide. The bacterium is stained pink.
A micrograph of Leptospira interrogans, a spirochete shaped bacterium, as seen with a scanning electron microscope. Several spiral shaped bacteria are seen overlapping each other. The bacteria are about 10 micrometers long and 0.25 micrometer wide.
A micrograph of Streptococcus pneumoniae, a coccus shaped bacterium, as seen under a light microscope. Several long chains of spherical bacteria are seen. Each bacterium is about 0.1 micrometer in diameter.
A micrograph of Streptococcus salivarius, a coccus shaped bacterium, as seen with a scanning electron microscope. Several long chains of spherical bacteria overlap each other. Each bacterium is about 1 micrometer in diameter.
FIGURE 2.6 ■Common shapes of bacteria.A, C, E. The shapes of most bacterial cells can be discerned with light microscopy (LM), but their subcellular structures and surface details cannot be seen. B, D, F. Surface detail is revealed by scanning electron microscopy (SEM). These SEM images are colorized to enhance clarity. DR. A. M. SIEGELMAN/VISUALS UNLIMITED, INC.DENNIS KUNKEL MICROSCOPY/SCIENCE SOURCEMICHAEL ABBEY/VISUALS UNLIMITEDDENNIS KUNKEL MICROSCOPY/SCIENCE SOURCEDR. A. M. SIEGELMAN/VISUALS UNLIMITED, INC.DENNIS KUNKEL MICROSCOPY/SCIENCE SOURCE